|
The Challenge of Understanding Northern Yellowstone Elk Dynamics after Wolf Reintroduction
by Daniel R. MacNulty, Daniel R. Stahler, C. Travis Wyman, Joel Ruprecht, & Douglas W. Smith
The status and trend of the northern Yellowstone elk herd has been an enduring conservation issue throughout the history of Yellowstone National Park. It is the largest of about seven migratory elk herds that graze the park's high-elevation meadows during summer. But unlike the other herds, the northern Yellowstone herd has a history of spending winter primarily within the park, ranging across the low-elevation grasslands and shrub steppes that fan out from the Yellowstone River and its tributaries along the park's northern border and adjacent areas of Montana. As the size of the northern herd has fluctuated over time, concerns have alternated between worries of too few and too many elk. Consensus about the appropriate size of the northern Yellowstone elk herd has been elusive.
This cycle of discontent originated in the late 19th century when concern focused on dwindling elk numbers due to market hunting and poaching (Houston 1982). Early 20th century protectionist policies, including the elimination of wolves and cougars, boosted elk numbers and stoked concern that the herd was too large. In response, park managers and hunters shot, trapped, and relocated tens of thousands of elk between 1920 and 1968, pushing the pendulum of public concern back toward concerns about too few elk. Then in 1969, the park implemented a policy of ecological process management known as natural regulation (Leopold et al. 1963), where elk numbers were allowed to fluctuate according to prevailing environmental conditions. Outside the park, the State of Montana used hunting to manage elk numbers. Except for the drought, fire, and severe winter of 1988–1989, conditions during 1968–1994 were generally favorable for elk survival and recruitment, and their numbers soared (figure 1). In turn, so did criticism that overabundant elk were destroying winter range vegetation.
A key outcome of this latter 20th century period, one that remains integral to understanding current elk dynamics, was a substantial increase in the distribution and abundance of elk wintering in the Yellowstone River valley outside the park. High elk densities inside the park, protection and restoration of winter range outside the park, and changes in the structure and timing of hunts in Montana more than doubled the winter distribution of elk north of the park (Lemke et al. 1998). It is unclear whether this shift represented a new condition or the return to a former one because historic records about the extent to which the northern herd wintered north of the park are ambiguous and debated (Houston 1982, Wagner 2006). Regardless, the expanded distribution of the northern herd into Montana raised concerns about overgrazing and agricultural damage on non-park lands. In 1976, the State of Montana lifted an 8-year ban on hunting migrant elk outside the park in December through February. In later years, the limited-permit late season hunt targeted mainly adult female elk with a goal of limiting numbers of elk wintering outside the park. During 1976–1995, the late season hunt removed an average of 965 total elk per year (range = 0-2,409 elk); whereas, the annual fall hunt removed 520 (range = 194- 2,728 elk; Lemke et al. 1998).
Consistent with expectations that hunting alone would not limit the size of the northern elk herd (Houston 1982, Mack and Singer 1993), a record high number of 19,045 elk were counted in January 1994 (figure 1). But it did not last. In December 1994, three months before the first set of reintroduced wolves exited their acclimation pens near the lower Lamar River, managers counted 2,254 fewer elk than during the previous winter for reasons not fully explained by harvest, since only 772 elk were removed during the preceding fall and late hunts (Lemke et al. 1998). And so began the latest major drop in northern Yellowstone elk numbers. By 2013, managers counted 3,915 elk, only 743 more than the herd's lowest count in 1968. Similar to previous declines, this one was met with widespread public consternation. Except this time wolves, not humans, received most of the blame.
When the policy of natural regulation was adopted to guide elk management in the park, predation was not considered essential to controlling elk numbers. Rather, food limitation alone was thought sufficient to limit the elk herd (Cole 1971, Houston 1976). Nevertheless, the policy's subsequent emphasis on maintenance and restoration of ecosystem processes paved the way for wolf reintroduction in 1995–1997. As a result, understanding the extent to which wolves are responsible for the latest decline in the northern elk herd is vital to gauging the consequences of a core prescription of natural regulation. It is also necessary to testing broadly important ideas about the ecological role of top predators. In particular, the hypothesis that wolves are ecosystem engineers that have suppressed elk herbivory and triggered large-scale recoveries of aspen and willow in northern Yellowstone (e.g., Ripple and Beschta 2012, Painter et al. 2015) assumes that wolves are a principle cause of the elk decline. However, scientific consensus about the role of wolves in driving the dynamics of the northern herd has yet to emerge, despite 20 years of research by numerous federal, state, and academic investigators.
An overarching reason for the impasse is that wolf reintroduction was neither a controlled nor replicated experiment. Political and financial constraints aside, such an experiment was impossible because there were no comparable elk herds living under similar environmental and management conditions. The northern Yellowstone herd was, and remains, a unique population. Also, numerous factors besides wolves affect elk population growth (e.g., summer precipitation, winter severity, and other predators including humans) and none were held constant. On the contrary, these factors varied enormously in the years after wolf reintroduction. Under these uncontrolled and unreplicated conditions, highly confident conclusions about cause and effect are difficult (perhaps impossible) to obtain. The challenge of inferring causation helps explain why the debate about the influence of wolves on northern Yellowstone elk dynamics is unsettled and why it will remain so for the foreseeable future.
In lieu of an experiment, the only tool scientists have to disentangle the cause(s) of the recent elk decline is long-term observation. This approach attempts to infer causation from strong correlation between annual measures of key system attributes (assuming these are known and measurable) across the observed range of variation. Spurious correlations can be avoided, or weakened, by collecting and integrating time series data on multiple expressions of the relationship of interest. For example, analysis of the correlation between elk population growth rate and wolf population size is strengthened by complimentary data on the relationship between elk calf recruitment and wolf predation rate.
A virtue of the northern Yellowstone ecosystem is it has been monitored longer and more intensively than most other ecosystems. As a result, many different time series data exist that are pertinent to understanding the forces that shape the dynamics of the northern elk herd. But there are obstacles with these data. First, the data are discontinuous. Financial and logistical constraints hinder faithful collection of annual data as well as limit some monitoring to short time periods. This leads to data gaps which obscure the link between cause and effect.
Second, the data are not necessarily accurate. Take for example the annual northern Yellowstone winter elk count, which has evolved over the last century from ground surveys taken over multiple days to aerial surveys conducted in a single day (Lemke et al. 1998). Modern aerial counts are known to be underestimates of true abundance (Houston 1982, Coughenour and Singer 1996, Singer et al. 1997, Eberhardt et al. 2007); but scientists, managers, and the public have mainly ignored this bias and interpreted the counts as estimates of true population size. Highlighting the danger of this approach, Singer and Garton (1994) estimated aerial surveys during 1986-1991 overlooked 9–51% of the northern elk herd, and the fraction of missed elk ranged from 9–30% in years with "good" sighting conditions to 35–51% in years with "poor" sighting conditions. This means ignoring annual changes in sightability can distort understanding of population trend. For example, counts during 1987 (17,007 elk) and 1988 (18,913 elk) suggested an increasing population, yet sightability-corrected counts for these years indicated a slight decrease (1987 = 23,350 elk; 1988 = 22,779 elk; Coughenour and Singer 1996). Fortunately, an outcome of the current elk decline is that researchers and managers have teamed up to build a statistical tool that will allow them to correct future elk counts for imperfect sightability, which will in turn strengthen inferences about the effects of wolves and other factors on elk population trend.
Despite uncertainty about the northern Yellowstone elk data, there is little doubt that wolves have contributed to the recent decline of the northern elk herd. What is in doubt is the size of that contribution. How much of the decline is due to wolves? The basic biology of wolves suggests that they have a modest influence on elk dynamics. The wolf has the bite force, body size, and cooperative behavior to kill a wide array of ungulates ranging from diminutive deer to one-ton bison (Mech et al. 2015). But it lacks the massive size, retractable claws, supinating muscular forelimbs, and specialized skull configuration (Peterson and Ciucci 2003) that would allow it to be a consistently high-success hunter of any one particular prey species.
Instead, the wolf is a consistently low-success hunter of a wide range of prey. Its strategy is to find the easy mark: a prey animal that is easily killed because of its small size, old age, poor health, or treacherous surroundings. The problem is that easy marks are generally rare and often inconspicuous. Wolves find their mark by relentlessly sifting through the available prey pool, testing prospective victims. Wolves cast a wide net and test many more prey than they actually kill. This is why the success of wolves hunting elk in northern Yellowstone has rarely exceeded 20% (Smith et al. 2000, Mech et al. 2001) and drops to less than 10% when only adult elk are considered (MacNulty et al. 2012).
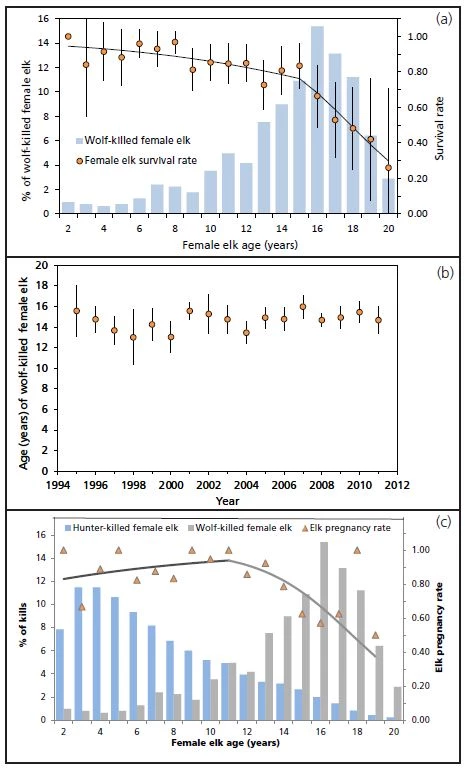
Selective hunting behavior of wolves determines the age distribution of prey they kill. Roughly half of elk killed by wolves in northern Yellowstone are calves, a pattern that has changed little since wolf reintroduction (Smith et al. 2004, Wright et al. 2006, Metz et al. 2012). Also unchanged has been the age distribution of the adult (more than 2 years-old) female elk they kill: 89% of 640 wolf-killed adult female elk in northern Yellowstone during 1995–2011 were more than 10-years old (figure 2a), and the annual mean age of these elk varied between 13- and 16-years-old (figure 2b). Nearly half (48%) of 606 wolf-killed elk documented in the Madison headwaters area of Yellowstone National Park during 1996–2007 were also calves (Becker et al. 2009). Older elk (10-13 years-old) also represented the largest overall share of adult females killed by wolves. However, the mean age of wolf-killed adult female elk was nearly 6 years younger in the Madison headwaters (9.1 years, 95% CI = 8.6, 9.7, n = 220) than in northern Yellowstone (14.7 years, 95% CI = 14.3, 15.0, n = 640). Extreme winter conditions and other factors contribute to a shorter lifespan and make elk more vulnerable to wolf predation in the Madison headwaters area ("Wolf Effects on Elk Inhabiting a High Risk Landscape: The Madison Headwaters Study," this issue). Selective wolf predation is important to the fate of the northern Yellowstone elk herd because it results in higher survival for the subset of elk that are rarely killed by wolves. This is evidenced by the relatively high mean annual survival (84-97%) of 2–to 8-year-old female elk in northern Yellowstone during 2000-2012 (figure 2a). Because this subset includes the most fertile females in the population (figure 2c), selective predation may reduce the impact of wolves on elk abundance (Wright et al. 2006, Eberhardt et al. 2007). On the other hand, selective predation also means wolves are major predators of elk calves, and calf survival may be the most important driver of elk population growth (Raithel et al. 2007). Thus, the effect of wolves on calf survival is arguably the single largest determinant of their role in the decline of the northern elk herd (Proffitt et al. 2014). But it is also one of the least understood aspects of wolf-elk interactions.
Existing information about the effect of wolves on calf survival in northern Yellowstone is not clear-cut. Long-term data on the composition of wolf-killed prey show that elk calves represent a large proportion of wolf-killed elk, particularly in summer (62%) and early winter (49%; Metz et al. 2012). Although suggestive, wolf-kill data do not measure calf survival per se. Barber-Meyer et al. (2008) provided a proper analysis of calf survival in northern Yellowstone by using radio-telemetry to track the fates of 151 newly born calves during 2003–2005 when wolf numbers peaked in northern Yellowstone (Cubaynes et al. 2014). They found that wolves accounted for only 14–17% of calf deaths and that overwinter calf survival was high (mean = 90%). It is likely the sample of calves entering each winter was too small (n = 12–16) to provide an unbiased estimate of overwinter survival. However, a comparable radio-telemetry study of northern Yellowstone calf survival conducted before wolf reintroduction (1987–1990) followed a larger sample of calves entering winter (n = 16–25) and found a similarly high rate of overwinter survival (mean = 86-94%) except in the severe post-fire winter of 1988-1989 (mean = 16%; Singer et al. 1997).
By contrast, summer survival rates of calves in 1987–1990 (mean = 65%; Singer et al. 1997) were more than twice that of those in 2003–2005 (mean = 29%; Barber-Meyer et al. 2008). Although at least some of the decrease was due to how the recent study defined the summer survival interval (capture date to October 31) to be two months longer than in the earlier study (capture date to August 31), it is notable that the proportion of calves killed by grizzly bears and black bears jumped from 23% (1987–1990;Singer et al. 1997) to as much as 60% (2003–2005;Barber-Meyer et al. 2008). This change aligns with an increase in the number of grizzly bears in the Greater Yellowstone Ecosystem during 1982-2007 (Kamath et al. 2015). These patterns would minimize the influence of wolves on calf survival, if not for the sheer number of elk calves among wolf-killed elk (Metz et al. 2012).
A similar discrepancy applies to cougars, which also commonly kill elk in northern Yellowstone. Like wolves, the composition of elk killed by cougars is dominated by calves (Ruth et al., in press). Moreover, the average total number of cougars inhabiting northern Yellowstone increased 76% from 1987–1993 to 1998–2004 (Ruth et al., in press). Yet, the proportion of cougar-killed radio-collared calves changed very little between 1987–1990 (1.5%; Singer et al. 1997) and 2003–2005 (2.6%; Barber-Meyer et al. 2008). Spatial mismatch between winter distributions of wolves, cougars, and radio-collared calves most likely explains why these predators killed so few radio-collared calves during 2003–2005 despite the prevalence of calves in their diets (Barber-Meyer et al. 2008).
As a result, questions persist about whether wolf, bear, and cougar predation adds to or replaces other sources of calf mortality, such as winter severity and other predators (Singer et al. 1997). On average, are wolves killing calves in northern Yellowstone that would otherwise survive their first year of life? A strong negative relationship between a proxy for calf survival (number of calves per 100 adult females counted in late winter; calf:cow ratios) and wolf population size (figure 3) is consistent with the hypothesis that wolves are an additive source of calf mortality. But inferring causation from this correlation is not foolproof. Calf:cow ratios are a composite of fecundity and calf survival, and may be confounded by changes in female age structure (Bonenfant et al. 2005). In addition, parallel changes in wolf abundance and other factors that affect calf survival, (e.g., bear/cougar abundance) confound assessment of a wolf effect.
Between the potential bias of analyzing calf:cow ratios and the high cost of radio-collaring and tracking calves, there are few, if any, good options for annually monitoring overwinter calf survival and the factors that affect it. In an effort to develop an alternative, researchers have started visually tracking the fates of calves observed at heel among about 70 radio-collared adult female elk that winter in northern Yellowstone. Although these observational data are also error-prone, they provide a valuable auxiliary dataset for assessing the validity of calf:cow ratios, as well as permit analyses of adult female reproductive success that account for the effects of individual-level factors such as age. Continued emphasis on assessing the effects of wolves on calf survival is an essential step toward greater understanding of the impact of wolves on the abundance of the northern Yellowstone elk herd.
Whereas debate about the magnitude of the effect of wolves on elk abundance is unresolved, there is a growing understanding that factors besides wolves contributed to the decline of the northern elk herd. Foremost among these are other predators, especially humans. In contrast to the age-selective predation patterns of wolves, cougars, and bears, human hunters participating in the northern Yellowstone late season hunt primarily killed the most fertile adult females (figure 2c). This likely represented a random sample of the female elk age distribution because the late hunt emphasized antlerless elk. By itself, regulated hunter harvest of young adult females is unlikely to reduce elk numbers. This is evidenced by substantial growth of the northern herd from 1976 to 1988 (figure 1) when hunters harvested large numbers of antlerless elk in the absence of much carnivore predation. And because hunters killed relatively few calves, high calf survival likely offset the removal of young adult females.
Elk calves enjoyed a large, perhaps unprecedented degree of protection from predation during the first 10–20 years of the natural regulation era. This began to change by the late 1980s when it became clear that a recovering grizzly bear population was increasingly preying on elk calves (French and French 1990). Growing cougar numbers and eventual wolf reintroduction increased predation pressure still further. By the early 2000s, the once predator-sparse environment of northern Yellowstone National Park was filled with record numbers of wolves, cougars, and grizzly bears preying on elk calves. Meanwhile, hunters continued to harvest substantial numbers of mainly young adult female elk during the late hunt. From 1995-2002, the late hunt annually removed between 940 and 2,465 total elk (figure 4). In 1997, severe winter conditions pushed many elk north of the park where they were exposed to hunter harvest. This resulted in the greatest number of elk harvested during the late season hunt (2,465 elk) since it was reinstated in 1976. And together with elk harvested during the preceding fall hunt, the total number of hunter-harvested elk during winter 1996–1997 represented the second largest removal of elk (3,320 animals) in the natural regulation era (figure 4). Record numbers of winter-killed elk suggest many harvested animals would have died of starvation had they avoided hunters. With continued declines in elk numbers observed during annual counts, the State of Montana reduced the number of late-hunt permits to less than 200 beginning in 2005 and suspended the hunt indefinitely following the 2009 season. While the fall season hunt continues, antlerless elk harvest has averaged less than 50 animals per year or less than 2% of the observed elk population since 2010 (Loveless 2015). The decline in hunting opportunity has fueled debate on the effects of predators on the northern herd, with the hunting public questioning the maintenance of high predator densities at the expense of hunting opportunity.
The decade following wolf reintroduction involved a level and pattern of predation on the northern herd it probably had not experienced since the market killing era (1872–1882) when wolves, cougars, and bears were probably still fairly abundant. The level of predation between 1923 and 1968 was also quite high, but this was mainly from humans (Houston 1982). As a result, the age of hunter-killed elk was not biased toward calves (e.g., Greer and Howe 1964) as it is with carnivore-killed elk. By contrast, the period between 1995 and 2005 involved a combination of carnivores killing calves and hunters killing young, fertile females. Wolf predation on old females may have also had a role, if diminished calf recruitment shifted the female age distribution toward older, more vulnerable age classes. Under conditions of intense predation across all ages of elk, it is difficult to imagine how the northern herd could have avoided a steep drop in abundance. Indeed, the mix of carnivore and human-caused mortality that defined this period may partly explain why the rate of decline after wolf reintroduction was greater than it was during 1923–1968 (figure 1) when humans were the only major predator. Declining ungulate abundance with increasing predator diversity has also been observed in moose and caribou systems (Gasaway et al. 1992, Peterson 2001). These studies suggest each additional predator species (i.e., wolves, grizzly bears, black bears, humans) results in a stepwise reduction in ungulate abundance. However, the dynamics and mechanics of this relationship are poorly understood. For example, little is known about how changes in the relative abundance of different predator species offsets (or exacerbates) the impact of predator diversity on ungulate abundance. In addition, it is unclear whether the combined effects of multiple predators on shared prey is the sum of their separate effects, or whether predators interact synergistically (or antagonistically) such that their combined impact is greater (or less) than the sum of their individual impacts.
The ability of grizzly bears to usurp wolf-killed elk (Ballard et al. 2003) suggests the potential for a synergistic effect; whereas, diminished cougar predation on elk calves in the presence of wolves and grizzly bears (Griffin et al. 2011) suggests a possible antagonistic effect. There is also the possibility that good forage conditions buffer elk against predation in systems with as many predators as Yellowstone (Griffin et al. 2011). Clearly, progress toward understanding the fate of the northern herd requires continued attention to northern Yellowstone as a multi-predator system.
A continued focus on the role of humans is also necessary. Cessation of the late hunt and reduced antlerless harvest during the general hunt in recent years provides a unique opportunity to assess whether adjusting human harvest can offset the impact of multiple carnivores on the abundance of the northern herd. Increased ungulate abundance in response to fewer predator species, including humans (Peterson 2001) together with evidence that human harvest has an overriding influence on adult female elk survival (Brodie et al. 2013) and elk population growth (Vucetich et al. 2005) suggests the northern herd may at least stabilize in the years ahead. If so, it will highlight how the fate of the northern herd is ultimately in the hands of humans, much as it has been since at least 1872.
The final actor in the northern Yellowstone saga that cannot go unmentioned is bison. A common refrain among those of us who were on the ground in northern Yellowstone during the late 1990s is that where we once saw herds of elk, we now see herds of bison. This has fueled speculation that bison are competing with elk and increasing bison numbers have contributed to the decrease in the northern elk herd following wolf reintroduction. This is an interesting reversal of perspective from the 1970s and 1980s, when the concern was about too many elk outcompeting bison and other ungulates. Studies during that period concluded that competition between elk and bison was minimal (Houston 1982, Singer and Norland 1994, Barmore 2003). Whether or not this still holds true is the subject of ongoing research.
No matter how much science tells us about what drives northern Yellowstone elk population dynamics, science alone is unlikely to resolve stakeholder concerns about too few or too many elk. This is because these concerns are less about science and more about competing visions of what northern Yellowstone should look like. What is indisputable is that the current version of the northern Yellowstone system (i.e., fewer elk wintering mainly outside the park, more bison wintering mainly inside the park, lower human harvests, high carnivore predation from multiple predators) is unlike any that has existed since managers conducted what was perhaps the first systematic count of the northern herd a century ago (Bailey 1916). How long this version lasts and what the next one may look like are fascinating questions. The answers will only be revealed if northern Yellowstone's many stakeholders continue to support long-term coordinated monitoring and assessment.



Literature Cited
Bailey, V. 1916. Letter to superindentent, Yellowstone National Park. Yellowstone National Park Heritage and Research Center, Gardiner, Montana, USA.
Ballard, W.B., L.N. Carbyn, and D.W. Smith. 2003. Wolf interactions with non-prey. Pages 259-271 in L.D. Mech and L. Boitani, editors. Wolves: behavior, ecology, and conservation. University of Chicago Press, Chicago, Illinois, USA.
Barber-Meyer, S.M., L.D. Mech, and P.J. White. 2008. Elk calf survival and mortality following wolf restoration to Yellowstone National Park. Wildlife Monographs 169:1–30.
Barmore, W.J. 2003. Ecology of ungulates and their winter range in northern Yellowstone National Park: research and synthesis, 1962-1970. National Park Service, Yellowstone National Park, Mammoth, Wyoming, USA.
Becker M.S., R.A. Garrott, P.J. White, C.N. Gower, E.J. Bergman, and R. Jaffe. 2009. Wolf prey selection in an elk-bison system: choice or circumstance? Pages 305-337 in R.A. Garrott, P.J. White, and F.G.R. Watson, editors. The ecology of large mammals in central Yellowstone: sixteen years of integrated field studies. Elsevier, New York, New York, USA.
Bonenfant, C., J.M. Gaillard, F. Klein, and J.L. Hamann. 2005. Can we use the young:female ratio to infer ungulate population dynamics? An empirical test using red deer Cervus elaphus as a model. Journal of Animal Ecology 42:361-370.
Brodie, J., H. Johnson, M. Mitchell, P. Zager, K. Proffitt, M. Hebblewhite, M. Kauffman, B. Johnson, J. Bissonette, C. Bishop, J. Gude, J. Herbert, K. Hersey, M. Hurley, P.M. Lukacs, S. Mc- Corquodale, E. McIntire, J. Nowak, H. Sawyer, D. Smith, and P.J. White. 2013. Relative influence of human harvest, carnivores, and weather on adult female elk survival across western North America. Journal of Applied Ecology 50:295–305.
Cole, G.F. 1971. An ecological rationale for the natural or artificial regulation of ungulates in parks. Transactions of the North American Wildlife Conference 36:417–425.
Coughenour, M.B., and F.J. Singer. 1996. Elk population processes in Yellowstone National Park under the policy of natural regulation. Ecological Applications 6:573–593.
Cubaynes S., D.R. MacNulty, D.R. Stahler, K.A. Quimby, D.W. Smith, and T. Coulson. 2014. Density-dependent intraspecific aggression regulates survival in northern Yellowstone wolves (Canis lupus). Journal of Animal Ecology 83:1344–1356.
Eberhardt, L.L., P.J. White, R.A. Garrott, and D.B. Houston. 2007. A seventy-year history of trends in Yellowstone's northern elk herd. Journal of Wildlife Management 71:594–602.
French, S.P., and M.G. French. 1990. Predatory behavior of grizzly bears feeding on elk calves in Yellowstone National Park, 1986-88. International Conference on Bear Research and Management 8:335–341.
Gasaway, W.C., R.D. Boertje, D.V. Grangaard, D.G. Kelleyhouse, R.O. Stephenson, and D.G. Larsen. 1992. The role of predation in limiting moose at low densities in Alaska and Yukon and implications for conservation. Wildlife Monographs no. 120.
Greer, K., and E. Howe. 1964. Winter weights of northern Yellowstone elk, 1961–62. Transactions of the North American Wildlife Conference 29:237–248.
Griffin, K.A., M. Hebblewhite, H.S. Robinson, P. Zager, S.M. Barber-Meyer, D. Christianson, S. Creel, N.C. Harris, M.A. Hurley, D.H. Jackson, B.K. Johnson, L.D. Mech, W.L. Myers, J.D. Raithel, M. Schlegel, B.L. Smith, C.G. White, and P.J. White. 2011. Neonatal mortality of elk driven by climate, predator phenology and predator community composition. Journal of Animal Ecology 80:1246–1257.
Houston, D.B. 1976. Research on ungulates in northern Yellowstone National Park. Pages 11–27 in Research in the parks. Transactions of the National Park Centennial Symposium, December 1971. National Park Service Symposium Series No. 1.
Houston, D.B. 1982. The northern Yellowstone elk herd. Macmillan, New York, New York, USA.
Kamath, P.L., M.A. Haroldson, G. Luikart, D. Paetkau, C. Whitman, and F.T. van Manen. 2015. Multiple estimates of effective population size for monitoring a long-lived vertebrate: an application to Yellowstone grizzly bears. Molecular Ecology 24:5507-5521.
Lemke, T.O., J.A. Mack, and D.B. Houston. 1998. Winter range expansion by the northern Yellowstone elk herd. Intermountain Journal of Sciences 4:1–9.
Leopold, A.S., S.A. Cain, D.M. Cottam, I.N. Gabrielson, and T.L. Kimball. 1963. Wildlife management in the national parks. Transactions of the North American Wildlife and Natural Resources Conference 28:28–45.
Loveless, K. 2015. Winter 2015 Hunting District 313 elk survey. Montana Fish, Wildlife and Parks. Bozeman, Montana, USA.
Mack, J.A., and F.J. Singer. 1993. Population models for elk, mule deer, and moose on Yellowstone's northern winter range. Pages 270–305 in R.S. Cooke, editor. Ecological issues on reintroducing wolves into Yellowstone National Park. National Park Service Scientific Monograph.
MacNulty, D.R., D.W. Smith, L.D. Mech, J.A. Vucetich, and C. Packer. 2012. Nonlinear effects of group size on the success of wolves hunting elk. Behavioral Ecology 23:75–82.
Mech, L.D., D.W. Smith, K.M. Murphy, and D.R. MacNulty. 2001. Winter severity and wolf predation on a formerly wolf-free elk herd. Journal of Wildlife Management 65:998–1003.
Mech, L.D., D.W. Smith, and D.R. MacNulty. 2015. Wolves on the hunt: the behavior of wolves hunting wild prey. University of Chicago Press, Chicago, Illinois, USA.
Metz, M.C., D.W. Smith, J.A. Vucetich, D.R. Stahler, and R.O. Peterson. 2012. Seasonal patterns of predation for gray wolves in the multi-prey system of Yellowstone National Park. Journal of Animal Ecology 81:553–563.
Painter, L.E., R.L. Beschta, E.J. Larsen, and W.J. Ripple 2015. Recovering aspen following changing elk dynamics in Yellowstone: evidence of a trophic cascade? Ecology 96:252–263
Peterson, R.O. 2001. Wolves as top carnivores: new faces in new places. Pages 151–160 in V.A. Sharpe, B.G. Norton, and S. Donnelley, editors. Wolves and human communities. Island Press, Washington, D.C., USA.
Peterson, R.O., and P. Ciucci. 2003. The wolf as carnivore. Pages 104–130 in L.D. Mech and L. Boitani, editors. Wolves: behavior, ecology, and conservation. University of Chicago Press, Chicago, Illinois, USA.
Proffitt, K.M., J.A. Cunningham, K.L. Hamline, and R.A. Garrott. 2014. Bottom-up and top-down influences on the pregnancy rates and recruitment of northern Yellowstone elk. Journal of Wildlife Management 78:1383–1393.
Raithel, J., M. Kauffman, and D. Pletscher. 2007. Impact of spatial and temporal variation in calf survival on the growth of elk populations. Journal of Wildlife Management 71:795–803.
Ripple, W.J., and R.L. Beschta. 2012. Trophic cascades in Yellowstone: the first 15 years after wolf reintroduction. Biological Conservation 145:205–213.
Ruth, T.K., P.C. Buotte, and M.G. Hornocker. In press. Yellowstone cougars: ecology before and during wolf reestablishment. University Press of Colorado, Boulder, Colorado, USA.
Singer, F.J., and J.E. Norland. 1994. Niche relationships within a guild of ungulate species in Yellowstone National Park, Wyoming. Canadian Journal of Zoology 72:1383–1384.
Singer, F.J., and E.O. Garton. 1994. Elk sightability model for the Super Cub. Pages 47-49 in J.W. Unworth, F.A. Leban, D.J. Leptich, E.O. Garton, and P. Zager, editors. Aerial survey: user's manual. Idaho Department of Fish and Game, Boise, Idaho, USA.
Singer, F.J., A. Harting, K.K. Symonds, and M.B. Coughenour. 1997. Density dependence, compensation, and environmental effects on elk calf mortality in Yellowstone National Park. Journal of Wildlife Management 61:12–25.
Smith D.W., L.D. Mech, M. Meagher, W.E. Clark, R. Jaffe, M.K. Phillips, and J.A. Mack. 2000. Wolf-bison interactions in Yellowstone National Park. Journal of Mammalogy 81:1128–1135.
Smith, D.W., T.D. Drummer, K.M. Murphy, D.S. Guernsey, S.B. Evans. 2004. Winter prey selection and estimation of wolf kill rates in Yellowstone National Park, 1995–2000. Journal of Wildlife Management 68:153–166.
Vucetich, J.A., D.W. Smith, and D.R. Stahler. 2005. Influence of harvest, climate and wolf predation on Yellowstone elk, 1961–2004. Oikos 111:259–270.
Wagner, F.H. 2006. Yellowstone's destabilized ecosystem: elk effects, science, and policy conflict. Oxford University Press, Oxford, UK.
Wright, G.J., R.O. Peterson, D.W. Smith, and T.O. Lemke. 2006. Selection of northern Yellowstone elk by gray wolves and hunters. Journal of Wildlife Management 70:1070–1078.
Dan MacNulty is a wildlife ecologist with expertise in the ecology of predator-prey interactions. He has studied wolf-ungulate interactions in Yellowstone National Park since wolves were reintroduced there in 1995. He started as a field technician monitoring wolves after they were released, conducted research on the behavioral ecology of wolf-ungulate interactions as a graduate student (MS and PhD, University of Minnesota) and post-doctoral researcher (Michigan Tech. University), and now leads an NSF-funded study of the effects of wolves on elk demography and movement as an assistant professor (Utah State University). His work has been published in a variety of journals, including Ecology Letters, Journal of Animal Ecology, and Behavioral Ecology and is featured in a new book entitled Wolves on the Hunt: The Behavior of Wolves Hunting Wild Prey co-authored with Dave Mech and Doug Smith and published by the University of Chicago Press. |
Last updated: September 27, 2024