|
Temporal Variation in Wolf Predation Dynamics in Yellowstone: Lessons Learned from Two Decades of Research
by Matthew C. Metz, Douglas W. Smith, Daniel R. Stahler, John A. Vucetich, & Rolf O. Peterson
Beginning with the pioneering work of Adolph Murie (1944) in Mount McKinley (now Denali National Park) in 1939-1941, ecologists have long been interested in evaluating the factors influencing wolf predation dynamics. Murie, who had just recently studied coyote ecology in Yellowstone National Park (YNP), was hired to assess wolves' relationship with Dall sheep. Through detecting and evaluating >800 Dall sheep skulls, Murie suggested wolves tended to kill vulnerable prey, an observation that has since been found to be a driving force in wolf predation dynamics (Mech and Peterson 2003). Because wolves are coursing predators who typically hunt large prey, selecting vulnerable individuals minimizes their risk of being injured while hunting. Murie's study provided the first glimpse into wolf-prey relationships, and many ecologists have spent significant time since trying to advance our understanding of wolf predation dynamics.
Among studies of predation, wolf-prey relationships are among the most well studied and best understood. In fact, the study of wolves and moose on Isle Royale National Park, which began in 1958 and continues today, is the longest running predator-prey study in the world. Since 1971, researchers have evaluated predation dynamics for wolves preying on moose, which are the sole ungulate (hoofed mammal) on Isle Royale. Central to the evaluation of how wolf predation influences moose population dynamics, has been collecting information about kill rate (kills per wolf per day) and predation rate (annual percent of moose population killed by wolves). This work has shown that the influence of wolf predation on moose population dynamics varies considerably over time (Peterson et al. 2014), highlighting that the influence of predation is not static but rather temporally dynamic.
Long-term studies like the one on Isle Royale are relatively unique. Wolf restoration in Yellowstone, however, provided a significant opportunity to conduct a similar long-term study in a much different ecological system. That is, while Isle Royale is characterized by its simplicity, YNP is best described by its complexity, as it is home to high densities of multiple large predators (e.g., wolves, cougars, grizzly bears) and eight different species of ungulates, including elk, bison, and mule deer. Moreover, Yellowstone is also affected by differing management strategies inside and outside of the park (e.g., human hunting). Currently, our research investigating wolf predation dynamics in YNP has been ongoing for two decades. Here, we will only discuss our work for a subset of packs that are intensively monitored and primarily live on the northern range of the park.
For many studies investigating wolf predation, kill rate estimates from winter provide the foundation. A common observation among these studies is that wolves kill more frequently as winter progresses, which has been primarily attributed to prey being easier to capture as snow depth increases (e.g., Huggard 1993, Post et al. 1999). Our work in YNP supports this previous research, as we observe kill rates are greater in late winter (March) than in early winter (mid-November to mid-December; Smith et al. 2004, Metz et al. 2012). Additionally, late winter kill rates are highest in years when winter is more severe (Mech et al. 2001; Yellowstone Wolf Project, unpublished data). However, in multi-prey systems, understanding how wolf predation influences prey population dynamics also requires knowing what prey species wolves are selecting. Historically, elk are the dominant species killed by northern range wolves (average 92% of wolf kills during a particular winter). Of note is that while the average was 95% in the first ten years, the average has declined to 88% in the last ten years.
Ending here, though, would provide an incomplete picture of how dramatically the northern range wolf-prey system has changed over the last two decades. Twenty years ago in the winter of 1995-1996, there were at least 15,000 elk and ~900 bison on the northern range; today, there are at least 5,000 elk and ~3,500 bison (figure 1). And although wolves rarely kill bison during winter (1.6% of wolf kills), they also welcome a free meal as bison make up 5.3% of all acquired carcasses (i.e., killed or scavenged). Moreover, bison comprised an average of 6% of the biomass acquired by wolves during winter over the first decade (1995-2004), but 20% over the last decade (2005- 2014; figure 1). This shift in the importance of bison in the diet of northern range wolves emphasizes the finding on Isle Royale (among others) that temporal variation in predator-prey dynamics is a critical, and likely universal, characteristic of large carnivore-prey systems.
This increased use of bison is also likely driven by there being fewer vulnerable elk within the current, less dense elk population. And although elk could be the perfectsized prey for wolves, wolves still incur risks while hunting them and are therefore selective about the type of elk they take. Specifically, during early winter when adult elk are less vulnerable, wolves typically select for calves. But as snow depth increases and nutritional conditions decline, wolves select for adult males during late winter (Smith et al. 2004, Metz et al. 2012). Additionally, when wolves prey on adult elk, they select for older adults (Wright et al. 2006). This selective nature of wolf predation should be even more prominent during other seasons of the year. Unfortunately, our understanding of large carnivore-prey dynamics had been primarily limited to winter because prey remains were difficult to consistently detect during snow-free periods.
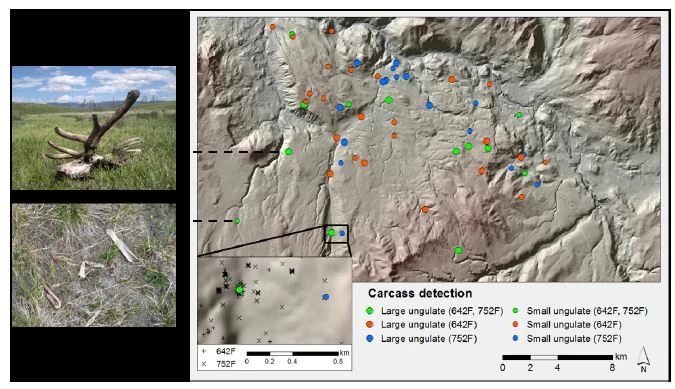
About a decade ago, a new window of opportunity was opening that would allow for a detailed look into how large carnivore predation dynamics differed throughout the year. Led by the pioneering work of Anderson and Lindzey (2003) studying cougar predation in southeast Wyoming, large carnivore-killed prey could be routinely located through searching carnivore GPS location "clusters" (i.e., spatially and temporally-related GPS collared carnivore locations; figure 2). With this new technique, precisely evaluating seasonal predation dynamics became possible. Our work in YNP has been at the forefront of this research, and we have examined wolf predation dynamics from May-July (encompassing elk calving season) through searching GPS clusters since 2004. Combining the data collected in spring (May) and summer (June and July) with data from winter, we have been able to evaluate precisely how wolf predation dynamics differ among the seasons of the year.
Ecologists had long expected seasonal differences in predation dynamics because of seasonal differences in prey availability and vulnerability. For the northern range wolf-prey system, ungulate migration affects prey availability, with most ungulates being more abundant within YNP during spring-fall. Through our work, we found the nutritional condition of ungulates killed by wolves is generally poorest during late winter and spring (figure 3). These seasonal differences in animal movement patterns and nutritional condition, along with the appearance of highly vulnerable neonate (newly born) ungulates during spring, drive seasonal variation in predation dynamics in YNP.
In comparison to winter, deer and bison are more prominent among wolf-killed prey in spring-summer. However, deer still make up a relatively small percent (9%) of all wolf kills found during this period. Bison (5%) also represent a small portion of wolf kills during spring-summer, although wolf-killed bison neonates are being increasingly detected in recent years (Yellowstone Wolf Project, unpublished data). Similar to winter, scavenging adult bison can also be an important food item for wolves in spring-summer. In particular, wolves sometimes scavenge adult female bison in the spring that likely die from birthing complications. Despite the use of these other species, elk (85%) is the dominant species killed by wolves during spring-summer. Among elk kills, wolves especially select for highly vulnerable neonate calves (64%) during summer. Wolves are one of many predators of elk calves in the predator-rich system of YNP; and although predation has been the leading cause of elk calf mortality since wolf restoration, bears are the dominant predator (~60% of deaths compared to ~15% for wolves; Barber-Meyer et al. 2008). Ultimately, the combined effects of predators (e.g., bears, wolves, cougars, coyotes) and climatic conditions play a critical role in elk calf survival rates (Griffin et al. 2011) and elk population growth rates (Raithel et al. 2007).
The most pronounced seasonal change in wolf predation dynamics is that kill rate differs throughout the year. Specifically, the number of prey that wolves kill per day peaks during summer, although kill rates of non-neonate elk (i.e., ≥6 months old) reach an annual minimum (figure 4a). Consequently, most wolf-killed prey during summer are neonates (62%) that provide little biomass. Most kills are neonates because adults are in better nutritional condition (figure 3) and are increasingly dangerous to hunt. Evidence for this increased risk is provided through the observation that wolves are most likely to be killed by an injury sustained from an ungulate during summer (figure 5). Our impression of seasonal variation in kill rate is also markedly different if we instead think about how much biomass wolves acquire. When doing so, the amount of food acquired by wolves is highest during late winter and spring when ungulate nutritional condition is poor (figure 3), and reaches its annual minimum during summer (figure 4b) when ungulate nutritional condition improves.
This seasonal pattern of biomass acquisition is driven by wolves being coursing predators whose own mortality risk varies throughout the year (figure 5). In comparison, cougars (ambush predators) display little seasonal variation in rates of biomass acquisition (Knopff et al. 2010). The differing rates that each of these carnivores acquires food throughout the year provides insight into their life history. That is, the seasonal curve for wolves (figure 4b) suggests wolves evolved to absorb the costs of reproduction (i.e., gestation and lactation from approximately mid-February to mid-June in YNP) during periods of the year when they acquire food in great excess of their energetic demands. Conversely, cougars can breed at any time of the year, although they tend to have young while neonate ungulates are being born (e.g., Elbroch et al. 2015). Ultimately, our work identifies summer as the limiting period of the year for wolves, and suggests that measuring kill rate during summer is required to understand how food acquisition affects wolf population dynamics.
The identification of summer as the limiting period for wolves is novel; yet the primary reason we began investigating predation dynamics 20 years ago was the same as Murie 75 years ago—to characterize the influence of wolves on prey population dynamics. This is a complicated topic with many factors that influence the strength of wolf predation on prey populations. Yet, for the simple system on Isle Royale, the percent of the moose population killed by wolves (i.e., predation rate) is a strong predictor of moose population growth rate, with the moose population being likely to decline when the predation rate exceeds 10% (Peterson et al. 2014). For Yellowstone, wolves' predation rate (% of elk population killed by wolves) is a poor predictor of elk population growth rate (Vucetich et al. 2011), likely because wolves are one of many factors (e.g., other predators, human harvest, climatic conditions) influencing elk survival. Nonetheless, wolf predation rate has increased in recent years (figure 6). This increase in the proportion of the elk population killed by wolves is due to the smaller elk population, rather than an increase in kill rate (i.e., how frequently wolves kill elk). If Isle Royale provides guidance to the consequences of temporal variation in predation rate, then the influence of wolf predation on the northern range elk population has likely increased in the most recent decade.
Our ability to estimate predation rate in YNP has been strengthened over the last decade because we have been able to precisely estimate seasonal wolf predation patterns. In doing so, we have gained a detailed understanding of many aspects of wolf predation. What we have learned so far indicates YNP's predator-prey system is temporally dynamic and much more complicated than just a wolf-elk system. Whether the first decade or the second is most representative of future wolf-prey relationships in YNP is unknown. Our findings, so far, provide significant insight into the annual cycle of wolf predation dynamics, and will allow for us to better understand why future conditions are similar or different to those witnessed during the last two decades.





Literature Cited
Anderson, C.R., and F.G. Lindzey. 2003. Estimating cougar predation rates from GPS location clusters. Journal of Wildlife Management 67:307-316.
Barber-Meyer, S.M., L.D. Mech, and P.J. White. 2008. Elk calf survival and mortality following wolf restoration to Yellowstone National Park. Wildlife Monographs 169:1-30.
Elbroch, L.M., P.E. Lendrum, P. Alexander, and H. Quigley. 2015. Cougar den site selection in the Southern Yellowstone Ecosystem. Mammal Research 60:89-96.
Griffin, K.A., M. Hebblewhite, H.S. Robinson, P. Zager, S.M. Barber-Meyer, D. Christianson, S.Creel, N.C. Harris, M.A. Hurley, D.H. Jackson, B.K. Johnson, W. L. Myers, J. D. Raithel, M. Schlegel, B.L. Smith, C. White, and P. J. White. 2011. Neonatal mortality of elk driven by climate, predator phenology and predator community composition. Journal of Animal Ecology 80:1246-1257.
Huggard, D.J. 1993. Effect of snow depth on predation and scavenging by gray wolves. Journal of Wildlife Management 57:382-388.
Knopff, K.K., A.A. Knopff, A. Kortello, and M.S. Boyce. 2010. Cougar kill rate and prey composition in a multiprey system. Journal of Wildlife Management 74:1435-1447.
Mech, L.D., and R.O. Peterson. 2003. Wolf-prey relations. Pages 131-160 in L.D. Mech and L. Boitani, editors. Wolves: behavior, ecology, and conservation. University of Chicago Press, Chicago, Illinois, USA.
Mech, L.D., D.W. Smith, K.M. Murphy, and D.R. MacNulty. 2001. Winter severity and wolf predation on a formerly wolf-free elk herd. Journal of Wildlife Management 65:998-1003.
Metz, M.C., D.W. Smith, J.A. Vucetich, D.R. Stahler, and R.O. Peterson. 2012. Seasonal patterns of predation for gray wolves in the multi-prey system of Yellowstone National Park. Journal of Animal Ecology 81:553-563.
Metz, M.C., J.A. Vucetich, D.W. Smith, D.R. Stahler, and R.O. Peterson. 2011. Effect of sociality and season on gray wolf (Canis lupus) foraging behavior: implications for estimating summer kill rate. PLoS ONE 6:e17332.
Murie, A. 1944. The wolves of Mount McKinley. U.S. Government Printing Office, Washington, D.C., USA.
Peterson, R.O., J.A. Vucetich, J.M. Bump, and D.W. Smith. 2014. Trophic cascades in a multi-causal world: Isle Royale and Yellowstone. Annual Review of Ecology, Evolution, and Systematics 45:325-345.
Post, E., R.O. Peterson, N.C. Stenseth, and B.E. McLaren. 1999. Ecosystem consequences of wolf behavioural response to climate. Nature 401:905-907.
Raithel, J.D., M.J. Kauffman, and D.H. Pletscher. 2007. Impact of spatial and temporal variation in calf survival on the growth of elk populations. Journal of Wildlife Management 71:795-803.
Smith, D.W., T.D. Drummer, K.M. Murphy, D.S. Guernsey, and S.B. Evans. 2004. Winter prey selection and estimation of wolf kill rates in Yellowstone National Park, 1995-2000. Journal of Wildlife Management 68:153-166.
Vucetich, J.A., M. Hebblewhite, D.W. Smith, and R.O. Peterson. 2011. Predicting prey population dynamics from kill rate, predation rate and predator-prey ratios in three wolf-ungulate systems. Journal of Animal Ecology 80:1236-1245.
Wright, G.J., R.O. Peterson, D.W. Smith, and T.O. Lemke. 2006. Selection of northern Yellowstone elk by gray wolves and hunters. Journal of Wildlife Management 70:1070-1078.
Matt Metz is a Research Associate with the Yellowstone Wolf Project. He earned his MS from Michigan Technological University while studying seasonal variation in wolf predation dynamics in northern Yellowstone. Currently, he is also a PhD student at the University of Montana, where his dissertation research focuses on evaluating how ungulate populations are affected by spatial heterogeneity in wolf predation dynamics. |
Last updated: July 6, 2016