|
FIRE ISLAND
Ecological Studies of the Sunken Forest, Fire Island National Seashore, New York NPS Scientific Monograph No. 7 |

|
CHAPTER 8:
BIOGEOCHEMICAL RELATIONSHIPS: INTRASYSTEM TRANSFERS
Weathering
In humid regions weathering of the soil minerals is most intense near the soil surface where organic matter concentrations, biological activity, water movement, expansion-contraction cycles, and wetting-drying cycles are greatest (Barshad 1964). The intensity of soil mineral weathering in podzol soils is usually calculated from comparison of the highly weathered A2 soil horizon with the unaltered parent material (Lutz and Chandler 1946). This method is not applicable to the Sunken Forest system since no significant differences (at P = 0.05) were found between the cation concentrations in the surface 15-cm layer and the 15-30-cm layer (Appendix III). This fact by itself would tend to indicate a low rate of weathering or a young age of parent material. The differences in the mean cation concentrations of the surface and subsurface layers amounted to 0.5-14.4 g/m2, based on the density of minerals in the surface layer. Assuming the age of the Sunken Forest ecosystem to be 250 years, these differences would indicate transfers of cations of the soil mineral compartment at a rate of less than 0.05 g/m2/year, clearly a negligible amount.
The history as well as the composition of the sediments comprising Fire Island would also suggest that the generation of cations from the soil-mineral compartment occurs at a slow rate. Sediments of both glacial and Long Island headlands origin have been subjected to chemical leaching by sea water. The sediments presently found in the Sunken Forest area are composed mainly of minerals resistant to further weathering (Clarke 1924). Thus transfers out of the soil-mineral compartment by weathering and transfers into the compartment, such as secondary mineral formation, are probably very small.
Uptake
The amount of cations in the primary production were calculated from the annual productivity of various tissues and their cation concentrations (Table 23, Appendices II, III). As such they are not accurate measures of total annual uptake of cations by the living biomass, rather these estimations represent the cations present in the production of a single point in time. The assumption is made that the cation concentrations in wood and bark laid down during the current year are approximately the same as in the bulk tissue sample analyzed. The total annual uptake by the vegetation is extremely difficult to estimate due to the rapid subannual transfers into and out of the organic compartment (Voigt 1960a; Witherspoon 1964; Waller and Olson 1967).
Although the uptake of nutrients is primarily by root tissues (Gauch 1957), the leaf and bark tissues also have the capacity to absorb nutrients (Tukey et al. 1952; Boynton 1954). The uptake of nutrients by foliar organs may be the primary nutritional pathway for epiphytes and lower plants with poorly developed roots (Tamm 1950, 1953, 1958; Wherry and Chapen 1928). Foliar absorption may also have an important role in the intrasystem cycling patterns, particularly in the uptake by subordinate vegetation of nutrients leached from higher in the canopy (Tukey and Mecklenburg 1964).
| K | (%) | Na | (%) | Ca | (%) | Mg | (%) | |
| Tree layer | ||||||||
| Fruits | 0.02 | * | * | * | ||||
| Current twigs + leaves | 3.29 | (75) | 0.99 | (56) | 2.92 | (63) | 1.05 | (63) |
| Branch wood + bark | 0.37 | (8) | 0.25 | (14) | 0.88 | (19) | 0.16 | (9) |
| Stem wood + bark | 0.17 | (4) | 0.09 | (5) | 0.31 | (6) | 0.07 | (4) |
| Roots | 0.51 | (12) | 0.44 | (25) | 0.52 | (11) | 0.39 | (23) |
| Total | 4.63 | 1.77 | 4.62 | 1.67 | ||||
| Shrub layer | ||||||||
| Current twigs + leaves | 0.08 | 0.02 | 0.11 | 0.03 | ||||
| Herb layer | ||||||||
| Shoots | 0.28 | 0.03 | 0.12 | 0.06 | ||||
| Lianas | ||||||||
| Smilax leaves | 0.38 | 0.04 | 0.07 | 0.12 | ||||
| Total † | 5.11 | 1.85 | 4.92 | 1.88 | ||||
† Exclusive of shrub branch, stem and root; herb roots; liana stems and roots. * = <0.01% | ||||||||
Foliar absorption of nutrients is probably a very important process in the maritime forest due to high atmospheric salt concentrations and high relative humidity in coastal areas. The overall mechanism of foliar absorption is ion exchange at the leaf surface, diffusion through the cuticle, and active uptake in the plasmalemma (Keppel 1967; Yamada et al. 1966; Jyung and Wittwer 1965; Wittwer et al. 1967). To be taken up through foliar and branch surfaces, salts must be in an aqueous phase. A transition from the solid phase of a salt to the liquid phase takes place at relative humidities of less than 100%, the transition humidity being dependent upon the hygroscopicity of the particular salt (Twomey 1953). Comparisons of the order of relative speed of foliar absorption of various salts and the phase-transition humidities of these salts indicate the most easily absorbed salts are saturated at the lowest relative humidities (Barinov and Ratner 1959; Twomey 1953). Therefore, foliar uptake, like root absorption, is a selective process regulated by both the physical-chemical environment and the biological nature of the absorbing tissues.
Selective uptake may lead to distributions of nutrients in the primary production that are vastly different from those in the pool of available nutrients. The amounts of calcium, magnesium, and sodium in the primary production are 1/6, 1/5, and 1/4 the amounts of these cations in the available nutrient compartment (Tables 22, 24). In contrast, the amounts of potassium in the primary production and the available nutrient compartment are about the same. This pattern may be the result of both the order of strengths of the adsorptions of these cations in the available nutrient compartment (Ca++ > Mg++ > K+ > Na+ and the selective uptake of cations by the roots and shoots of the living biomass, there being 2.5 times more potassium and calcium than sodium and magnesium in primary production.
The distributions of cations in the primary production change throughout the growing season with a rapid accumulation of nutrients in developing tissues, sometimes at the apparent expense of other tissues (Tamm 1951; Mitchell 1936). The nutrient levels in the leaves become stable during the middle of the growing season when the samples were collected at Fire Island (Stenlid 1958). With the onset of dormancy and leaf fall, amounts of biologically mobile elements such as potassium, phosphorus, and nitrogen are retranslocated into perennial tissues, while immobile elements such as calcium and silicon remain in the senescent leaves as they fall (Sampson and Shamish 1935; Mitchell 1936).
While primary production in the Sunken Forest is only 6% of the total biomass dry weight, it accounts for at least 14% of the potassium, 8% of the sodium, 11% of the calcium, and 11% of the magnesium in the total living biomass (Tables 22, 24). Similar to the distribution of cations in the biomass, the tree layer accounts for the vast majority (85-95%) of the 5.1 g/m2 potassium, 1.8 g/m2 sodium, 4.9 g/m2 calcium, and 1.9 g/m2 magnesium in the primary production (Table 24).
| K | Na | Ca | Mg | |
| Weathering | * | 0.06 | 0.04 | 0.01 |
| Leaching | ||||
| Gross through fall | 3.29 | 14.45 | 2.24 | 2.30 |
| Gross stemflow | 0.75 | 4.14 | 0.59 | 0.53 |
| Gross total leaching | 4.04 | 18.59 | 2.83 | 2.83 |
| Litter fall | 0.87 | 0.69 | 6.11 | 1.35 |
| Uptake (production) Tree layer | ||||
| Ilex opaca | 2.92 | 1.12 | 2.12 | 1.12 |
| Sassafras albidum | 0.31 | 0.08 | 0.23 | 0.06 |
| Amelanchier canadensis | 0.95 | 0.52 | 2.11 | 0.44 |
| Other species | 0.18 | 0.05 | 0.16 | 0.05 |
| &mbsp; Tree total | 4.36 | 1.77 | 4.62 | 1.67 |
| Shrub layer µ | 0.09 | 0.02 | 0.11 | 0.03 |
| Herb layer µ | 0.28 | 0.02 | 0.12 | 0.06 |
| Lianas µ † | 0.38 | 0.04 | 0.07 | 0.12 |
| Total | 5.11 | 1.85 | 4.92 | 1.88 |
µMinimum estimates based on current twigs and leaves only. † Above ground Smilax only. * < 0.005 g/m2. Based on differences in cation concentrations of soil layer minerals (not significant at P = 0.05). | ||||
Annual tissues (leaves) contain greater amounts of cations than the perennial tissues (branches, bark, and wood) concurrently produced. Because of their high cation concentrations, current twigs plus leaves account for 56-75% of the cations in the tree-layer production. Branch wood plus bark comprise between 8 and 19% of the cations in production, while stem wood plus bark account for only 4-6%. Root tissues contained 11% of the calcium and 12% of the potassium in production, but 23-25% of the sodium and magnesium. The uptake of nutrients by the annual tissues will determine to a great extent the patterns of intrasystem cycling through the subsequent return of nutrients to the available pool through leaching and decomposition.
In virtually all forested ecosystems, there are greater amounts of nutrients in the production of annual tissues that can be readily cycled than in the annual increments of perennial tissues (Rodin and Bazilevich 1967; Rennie 1955; Ovington 1962, 1965, 1968). The sustained metabolism of forest ecosystems is dependent upon the rapid recirculation of the relatively limited capital of available nutrients.
Decomposition
Cations are transferred from the organic compartment to the available nutrient compartment either directly by leaching or indirectly through decompositional pathways involving secondary production within the organic compartment (Bormann and Likens 1967). While the total amounts of cations leached from the above-ground biomass can be measured, it is extremely difficult to separate the annual and subannual components of this transfer (Remezov 1959). The amounts of cations leached or exuded from root systems are largely unknown but potentially are an important transfer (Woods and Brock 1964; Smith 1969).
Similarly, the organic debris falling from the above-ground biomass is easily measured, but there is no convenient method of estimating the turnover of root tissues. The importance of root losses in forested ecosystems was emphasized by Orlov (1959), who estimated the annual turnover of roots <0.5 mm in diameter for 25- and 50-year-old fir stands to be 200 g/m2 and 60 g/m2, respectively.
The role of secondary producers in the Sunken Forest, although not measured directly, is assumed to be very important. As previously mentioned, there are large populations of fruit-eating birds (towhees, catbirds, thrashers, robins), earthworms, and isopods in the forest. The consumption of foliar tissues by phytophagous organisms during the period of this study was estimated to be only 0.3-1.2% of the leaves produced by the dominant tree species, a level which is generally lower than in other forested ecosystems (Appendix II) (Whittaker and Woodwell 1968). The breakdown of leaf tissue, therefore, appears to be mainly through detrital food chains rather than by consumption of living tissues. Organisms such as isopods which are associated with detritus consumption generally have low assimilation efficiencies and therefore have higher rates of substrate consumption and turnover compared to organisms feeding on living material (Reichle 1967). The main role of invertebrates feeding on forest litter appears to be the physical breakdown of litter rather than chemical alteration of the substrate (White 1968; Edwards et al. 1970).
Methods: Litter Analysis
The cation composition of the litter fall, along with the precipitation falling through the Sunken Forest canopy (throughfall), was sampled using the design of Wilm (1946), incorporating the statistical modifications of Rieley et al. (1969): A stationary collector (X) measures variation between collection periods (through time) and a series of collectors moved to new randomly selected locations (Yj) measure variation within collection periods (through space). The expected value for a given collection period (Ei) is calculated from the regression
Ei = cXi
where the least squares estimator of c is
c = 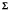 ij XiYij/kiX2i
ij XiYij/kiX2i
(Rieley et al. 1969).
The 20 X 30-m ecosystem analysis plot was subdivided into three parallel 20 X 10-m sections, each having one stationary and two moving collectors. The collectors consisted of a 20.4-cm diameter sheet-metal funnel coated with acrylic paint which fit onto a 2-gallon polyethylene bottle. At roughly weekly intervals between 6 October 1958 and 2 October 1969 (Appendix IV), collections were made and clean (acid washed, distilled water rinsed) polyethylene bottles were installed for the litter-throughfall collectors. The litter material was filtered from the throughfall with tarred Whatman No. 40 ashless filter paper. The litter fraction was dried at 85°C, weighed, and prepared for cation analysis following the procedures of Likens and Bormann (1970). The filtered throughfall fraction was measured volumetrically and retained for cation analysis. Since the lengths of collection periods varied, data are expressed on an amount per day basis and represent the means of the three 20 X 10-m units within the plot. The percent standard errors of the mean for the litter cation analysis were: dry matterplusmn;12% Kplusmn;12% Naplusmn;16% Caplusmn;14% and Mg±14%.
Results and Discussion
The estimated 489 ± 59 g/m2/year of litter fall from the litter-throughfall collectors is in good agreement with the estimates of 557 ± 55 g/m2/year from the litter baskets. The seasonal distribution of total litter fall from both methods of litter collection shows the same pattern of autumnal and vernal peaks (Fig. 28, Appendix IV).
The seasonal distribution patterns of cations in the litter fall generally reflect the trends in litter dry weight deposition (Figs. 28, 43). Sodium, magnesium, and calcium all have distinct autumnal and vernal peaks associated with leaf fall phenology and a peak in February associated with a severe winter storm. The potassium deposition in litter fall shows the distinct autumnal and February peaks but the vernal peak is not as pronounced as for the deposition of other cations. A possible explanation of the potassium distribution pattern may be that in Ilex opaca greater amounts of potassium than other cations are being retranslocated into perennial tissues from leaves that are undergoing senescence. The differences in autumnal and vernal deposition patterns would also suggest that the selective retranslocation of potassium is not as great in Sassafras and Amelanchier as in Ilex.
The transfer of litter represents the movement of 17% of the potassium, 37% of the sodium, 72% of the magnesium, and 124% of the calcium in the primary production to organic debris (Table 24). The anomaly of 1.24 times more calcium in the litter fall than in the primary production undoubtedly stems from the mid-July sampling of leaf tissues for primary production estimates. The amounts of calcium as well as magnesium in Sassafras and Amelanchier leaves would be expected to increase until autumn leaf fall, and in Ilex opaca these cations probably increase until leaf fall the following spring (Mitchell 1936; Lutz and Chandler 1946).
The transfer of cations from organic debris in the form of litter to the available nutrient compartment appears to be a rapid process. Not only does there appear to be a rapid physical breakdown of freshly fallen litter in the Sunken Forest, but leaching of the litter by rainwater undoubtedly hastens the transfer from the organic debris to the available nutrient compartment. The release of sodium and potassium from decomposing litter is usually at a faster rate than the release of calcium, magnesium, or phosphorus (Attiwill 1968). The amount of calcium relative to other cations is greater in the annual litter fall than in the organic debris as a whole and greater in the organic debris than in the available nutrient compartment (Tables 22, 24). This distribution pattern is the result of the more rapid leaching, mineralization, or transfer of potassium, sodium, and magnesium from the fresh litter and the subsequent retention of these cations by the lower humus layers (Burges 1967; Attiwill 1968).
|
| Fig. 43. Amounts of cations in litter fall (mg/m2/day). (click on image for an enlargement in a new window) |
Leaching
Methods
One form of transfer of cations from the organic compartment to the available nutrient pool takes place by throughfall, which is precipitation falling directly to the ground from the canopy; and by stemflow, the flow of precipitation down tree stems (Helvey and Patrick 1965). The leaching collections are actually gross stemflow and gross throughfall which also contains cations from salt spray and precipitation inputs. Collections of stemflow were made concurrently with the collections of throughfall previously described.
Stemflow was sampled by stratifying Ilex, Amelanchier, and Sassafras populations into three diameter-size classes and then randomly selecting one sample tree in each size class for each species. Aluminum stemflow collars were attached to each sample tree with steel strapping and were caulked with an inert compound. The collars were fitted with stainless steel screens over the effluent duct and the entire collar was then coated with an acrylic paint. A length of tygon tubing connected the collar to polyethylene collection tanks which were connected in tandem. The tanks, which remained in the forest for the duration of the stemflow collections, were painted with black paint to reduce algal growth.
Stemflow was measured volumetrically in the field for each collection period and was subsampled for cation analysis. However, stemflow collections were not made between 7 December 1968 and 27 March 1969, since precipitation in the form of snow and freezing of the collection tanks were frequent occurrences during this period.
The calculations of amounts of cations in the throughfall on a land area basis followed the methods used for cation analysis of litter fall (Wilm 1946; Rieley et al. 1969). The percent standard errors of the means for the throughfall were: amount of throughfall ±9% K ±9% Na ±8%, Ca ±7%, and Mg ±8%. Stemflow was converted to a land area basis by multiplying the mean ratio of stemflow:branch surface area of the three sample trees of each species by the total branch surface area of the species. The stemflow:branch surface area method was adopted due to the high variability of the sample trees and the morphological peculiarities of Sunken Forest trees which precluded the use of regressions on stem diameter. Regressions of stemflow on tree stem diameter or basal area gave multiple correlation coefficients of 0.81-0.88 while the r's for the method adopted were 0.95-0.99.
Results and Discussion
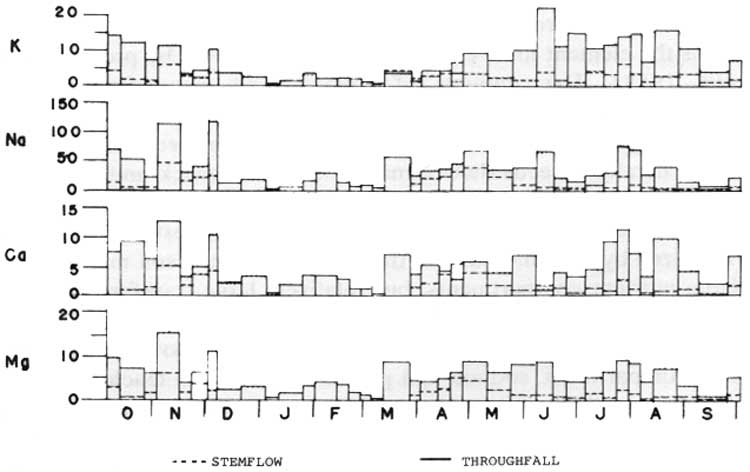
The 108.8 cm precipitation reaching the ground surface in the forest per year is equivalent to 93.6% of the 116.3 cm falling in the open. The 91.2 cm/year of throughfall accounts for 84% of the precipitation under the canopy, while 17.6 cm/year stemflow is only 16%. Although compared to the throughfall the amounts of stemflow are relatively small, they may be a particularly important component in the cycling of nutrients since the water flowing down the stems tends to follow the root systems into the soil (Voigt 1960b). Furthermore, the stemflow has higher cation concentrations than the throughfall (Table 26).
The seasonal distribution of amounts of cations leached from the Sunken Forest canopy exhibit a pattern of greater amounts in March through November than in December through February (Fig. 44). This pattern is undoubtedly due in part to the phenology of the dominant vegetation. The metabolic activity of the Sunken Forest is probably at a minimum between December and March, at which time the leaching transfers are also at a minimum level. The increases in leaching in the spring are concurrent with the onset of bud bursting, flowering, leafing out, and Ilex leaf fall, all of which are processes associated with the translocation of nutrients (Stenlid 1958). The amounts of cations leached from the canopy rapidly declines with the completion of autumnal leaf fall.
|
| Fig. 44. Amounts of cations in gross leaching (mg/m2/day). (click on image for an enlargement in a new window) |
Concurrent with the phenological changes in the forest vegetation are changes in the patterns of prevailing winds which undoubtedly affect the amounts of salt-spray aerosols impacted on the vegetative surfaces. During the spring, summer, and autumn, the prevailing winds are from the south and west, while during the winter the winds are from the northern compass quadrant (Figs. 9, 10). The onshore winds during the growing season generate more salt-spray aerosols than the northern winter winds blowing across the Great South Bay toward the forest. Thus the seasonal differences in gross leaching may be partially due to nutrient input patterns as well as the transfers of cations from the organic compartment to the available nutrient compartment.
The amounts of cations in the precipitation reaching the ground surface in the Sunken Forest (gross leaching) represent cation inputs in the forms of aerosol impaction and precipitation, plus cations leached from the organic compartment minus cations taken up by foliar and woody tissues. Although the cations in the precipitation can be corrected for, it is difficult to separate the other components in the gross leaching.
Sodium is by far the most abundant cation in the throughfall and stemflow (Table 24). Much of the sodium and probably much of the magnesium are salt-spray aerosols washed off the vegetative surfaces by the precipitation. Potassium, the second most abundant cation leached from the Sunken Forest canopy, is an extremely mobile cation which usually is the element most easily removed from leaves by precipitation (Stenlid 1958). The amounts of calcium and magnesium in the throughfall and the stemflow are about equal in the Sunken Forest, although in noncoastal forests, the amount of calcium leached from the canopy generally exceeds that of magnesium (Madgwick and Ovington 1959; Nye 1961; Attiwill 1966; Art 1974).
The circulation of cations within the Sunken Forest ecosystem is characterized by little measurable transfer between the soil mineral and available nutrient compartments, but relatively large transfers between the available nutrient and organic compartments. Cations have differing modes of transfer from the organic compartment to the available nutrient compartment, sodium and potassium largely in leaching, calcium largely via litter fall, and magnesium an intermediate situation (Table 24).
The amounts of cations held in the net primary productivity relative to the circulation of cations in leaching and litter fall (K 5.1:4.9; Na 1.8:19.3; Ca 4.9:8.9; Mg 1.9:4.2) may indicate the tightness of the cycles of various cations. Sodium, which is abundant in sea water and not an essential plant nutrient, is transferred through the system by leaching and litter fall in an amount seven times greater than in the primary production. Twice as much calcium and magnesium but about the same amount of potassium circulate by leaching and litter fall as are held in the net primary production. This suggests that potassium, which may be translocated to and from leaf tissues and perennial tissues, may be cycling within the living biomass to a greater extent than calcium or magnesium. This view is also supported by leaching experiments which suggested the movement of basipetally immobile nutrients such as calcium and magnesium may be external to plants (Tukey and Mecklenburg 1964).
| <<< Previous | <<< Contents >>> | Next >>> |
chap8.htm
Last Updated: 21-Oct-2005