|
MOUNT RAINIER
The Forest Communities of Mount Rainier National Park |

|
CHAPTER 7:
FOREST DYNAMICS
Change occurs constantly in forest landscapes. Disturbances and the ecological characteristics of the various tree species are responsible for most of this change. There is a continuum of disturbances, from the large, catastrophic event that destroys all or most of the old forest, to small-scale disturbances, such as an individual windthrown tree, that create much of the patchiness within established stands. Tree-species characteristics, such as their seeding habits and ability to tolerate shade, will determine how forest composition will evolve in response to these disturbance events.
In this chapter we will briefly review what is known about forest disturbance and succession in Mount Rainier National Park. We begin with considerations of large-scale disturbances, such as wildfire and volcanic events. Successional relationships of tree species and rates of forest recovery are considered next. The chapter concludes on the topic of tree mortality and patterns of small-scale disturbance in forests at Mount Rainier.
Large-Scale Disturbances
Catastophic disturbances are events that destroy most or all of the preceding forest, thereby creating conditions for establishment of a new forest stand. These infrequent events "reset" the forest to the beginning of a new successional sequence and open habitat for pioneer species. Succession is initiated with herb- and shrub-dominated communities under conditions of little or no forest canopy. New trees gradually become established, the tree canopy closes, and the forest then progresses through a series of structural and compositional changes. During stand development, species of lower shade tolerance (such as Pseudotsuga nnenziesii and Abies procera) are gradually replaced by more shade-tolerant tree species (such as Abies annabilis and Tsuga heterophylla).
At Mount Rainier National Park, catastrophic events have been of primary importance in creating the basic mosaic of forest patches of varying age and composition. Agents of forest destruction in the Park include wildfire, wind, volcanic events (especially lahars within recent times), snow avalanches, and floods. The frequency and distribution of many such events during the last millenium has been documented at Mount Rainier (Hemstrom and Franklin 1982).
Wildfire is, by far, the most important agent of forest catastrophe at Mount Rainier National Park, followed by snow avalanches and debris flows (lahars) (Hemstrom and Franklin 1982). Wildfires have occurred on approximately 90 percent of the existing stands, avalanches on 7 percent, and lahars on 2 percent. Small tracts of both blowdown and flooded forest are also present. While high-velocity winds can destroy extensive forested areas in the Pacific Northwest, as during the Columbus Day windstorm of 1982, wind generally appears to function as an agent of small-scale, within-stand disturbance at Mount Rainier National Park. Exceptions may be found along Park boundaries, where forests are exposed to the full force of winds by clearcutting on adjacent lands.
Fourteen major fire events have been documented since 1230 A.D. (Table 17 and Plate 2). The largest and oldest disturbance identified was a widespread fire episode in 1230 A.D. that affected about 25,000 ha of forest land in all four quarters of the Park. Stands of this 750-year age class still dominate much of the Ohanapecosh River drainage. Other episodes that affected approximately 13,000 ha of forest occurred in 1403 A.D., 1503 A.D., and 1628 A.D. The 350-year-old stands that originated following the 1628 fire are the most extensive age class of forest currently existing in Mount Rainier National Park; such forests dominate the upper slopes in the Carbon, Mowich, Puyallup, and, to a lesser extent, the Nisqually River drainages. One or more fires in 1703 A.D. resulted in the establishment of 275-year-old stands in the White and Ohanapecosh River drainages; stands of this age characterize the Ohanapecosh Campground and surrounding areas. Extensive fires that occurred in the mid to late 1800s may have been ignited by European settlers who entered the region at that time. Large portions of the Cowlitz drainage burned in 1856 and again in 1885, and the White River was subjected to a major fire in 1858. Large areas at moderate to high elevations within both of these drainages are still sparsely forested.
Table 17. Major fires, their correspondence to periods of drought, and the present and reconstructed original extent of resulting seral forests at Mount Rainier National Park (after Hemstrom and Franklin 1982).
| Episode date (A.D.) |
Present forested area |
Reconstructed forested area |
Drought period | ||
| Hectares | Percent of total |
Hectares | Percent of total | ||
| 1230 | 6,265 | 12 | 25,000 | 47 | |
| 1303 | 1,450 | 3 | 6,000 | 11 | 1290-952 |
| 1403 | 6,910 | 13 | 13,700 | 26 | 1406-132 |
| 1503 | 4,700 | 9 | 13,680 | 26 | 1477-902 |
| 1628 | 11,060 | 21 | 12,900 | 24 | 1627-332 |
| 1688 | 1,200 | 2 | 4,410 | 8 | 16863 |
| 1703 | 2,700 | 5 | 5,140 | 10 | 17003 |
| 1803 | 2,230 | 4 | 2,230 | 4 | 18013 |
| 1825 | 2,400 | 5 | 2,480 | 5 | 1823-263 |
| 1856 | 490 | 1 | 2,800 | 5 | 18563 |
| 1858 | 3,040 | 6 | 3,700 | 7 | 18563 |
| 1872 | 600 | 1 | 600 | 1 | 1869-732 |
| 1886 | 3,800 | 7 | 4,280 | 8 | 18883 |
| 1934 | 770 | 1 | 770 | 1 | 1917-362 |
1No climatic reconstructions available for dates before 1250 A.D. 2Keen (1937) first- or second-magnitude drought. 3Blasing and Fritts (1976) abnormally dry winter. | |||||
Fire frequency appears to be a complex issue based on the fire history reconstruction (Hemstrom and Franklin 1982). Natural fire rotation, defined as the time necessary for fires with a given frequency to burn over and reproduce an area the size of the study area, is about 465 years for the pre-European era at Mount Rainier. During early settlement, the fire rotation dropped to 226 years, perhaps because of human activity. Climatic influences appear important in creating conditions for catastrophic fire, however. Correspondence between drought events, as reconstructed from tree-ring chronologies, and fire episodes is striking (Table 17). Hence, major fires may be dependent upon unusual weather conditions and not reflect an internally induced cycle of forest susceptibility to burning.
Fire occurrence varies with topographic position, Park drainage, and habitat type (Hemstrom 1982, Hemstrom and Franklin 1982). Alluvial terraces, valley bottoms, and protected north-facing slopes have burned less frequently and are often occupied by old stands. Furthermore. nearly every major river valley contains a streamside old-growth corridor. Based on a study of fire-boundary locations within the Park, valley bottoms are, along with ridgetops, the most important natural fire-breaks (Hemstrom 1982). Contrasts in forest ages between north- and south-facing slopes also occur in almost every major drainage. Within the Park the White, Cowlitz, and Nisqually River drainages have burned the most frequently. In the case of the Cowlitz and Nisqually, this may be due to a generally south aspect and lack of topographic and vegetative barriers that might prevent fires from burning into the Park. Higher fire frequency is to be expected in the relatively dry White River drainage; natural fire rotation is 324 years for White River compared to 438 years for the Nisqually drainage (Hemstrom 1982).
Fire frequency varies rather drastically with habitat type (Table 18) (Hemstrom 1982). Much of this variation is correlated with topographic position. For example, several of the types with high fire frequency are characteristic of south slopes. Habitat types with low fire frequency are typically moist, valley-bottom sites (e.g., ABAM/OPHO) or cold, wet subalpine habitats (e.g., ABAM/RHAL).
Snow avalanches and lahars are other major forest disturbances that have been identified at Mount Rainier (Hemstrom and Franklin 1982). Approximately half of the 4270 ha of avalanche tracks are found in the White and Cowlitz River drainages. In general, large areas of snow avalanches are associated with recent burns; for example, large, complex tracks dissect the 1886 Cowlitz burn and the 1858 Crystal Mountain burn. Lahar-originated forests cover fairly small areas but occur in all of the major river drainages. The 1947 Kautz Creek lahar provides one of the most recent and obvious examples of a regenerating debris flow; young forests of Alnus rubra and conifers are typical of the deposits.
Table 18. Fire frequency1 (FF) and natural fire rotation2 (NFR) by habitat type for Mount Rainier National Park (excluding Carbon and Puyallup River drainages) (from Hemstrom 1982).
| Habitat or community type | NFR | FF | Rank |
| Abies lasiocarpa/Valeriana sitchensis | 275 | .0035 | 1 |
| Abies amabilis/Berberis nervosa | 295 | .0034 | 2 |
| Tsuga heterophylla/Achlys triphylla | 308 | .0033 | 3 |
| Abies amabilis and Tsuga heterophylla/Gaultheria shallon | 313 | .0032 | 4 |
| Abies amabilis/Xerophyllum tenax | 323 | .0031 | 5 |
| Abies amabilis/Menziesia ferruginea | 343 | .0029 | 6 |
| Abies amabilis/Rubus lasiococcus/Rubus lasiococcus phase | 367 | .0027 | 7 |
| Abies amabilis/Tiarella unifoliata | 426 | .0023 | 8 |
| Tsuga heterophylla/Polystichum munitum | 435 | .0023 | 9 |
| Abies amabilis/Vaccinium alaskaense and Chamaecyparis nootkatensis/Vaccinium ovalifolium | 474 | .0021 | 10 |
| Abies amabilis/Rhododendron albiflorum | 478 | .0021 | 11 |
| Abies amabilis/Oplopanax horridum | 535 | .0019 | 12 |
| Abies amabilis/Erythronium montanum | 616 | .0016 | 13 |
1Burned hectares per hectare of habitat type per year for fires over 100 ha. 2The time required to burn an area equal to the total area of each habitat type given its burn rate. | |||
Despite extensive catastrophic disturbances, forests at several locations around Mount Rainier have survived for over 1,000 years with no evidence of disturbance (Plate 2). These ancient forests grow in protected valley bottoms throughout the Park and on slopes in the Ipsut and Cataract Creek drainages, tributaries of Carbon River. Except for avalanches and a small burn in the upper reaches of the valley, the forests of the Ipsut Creek valley apparently have been free of major disturbances for over 1,200 years based on estimated ages of the large Chamaecyparis nootkatensis found there. Forests along Cataract Creek appear similar to those of Ipsut Creek but lack the Chamaecyparis; early seral trees are absent, and there is no evidence of catastrophic disturbance in the lower valley for well over 1,000 years. Other stands estimated as being over 1,000 years of age are found near the confluence of the Ohanapecosh River and Chinook Creek, in the upper Cowlitz River basin, and near Cougar Rock Campground in the Nisqually River drainage. The latter stand appears to be the first generation of forest on a portion of the Paradise lahar assemblage (Crandell 1971); trees have grown very slowly on this nutrient-poor substrate.
Successional Sequences
Occurrence and successional roles of the major tree species at Mount Rainier vary with moisture and temperature conditions (Table 19). The most shade-tolerant species occurring on a habitat type are usually considered to be the potential climax species, i.e., capable of maintaining themselves on the site in the absence of major disturbance. Tree species that are intolerant of shade and require a major disturbance for reproduction are considered to be seral. However, it is important to note that climax species often reproduce immediately following a disturbance and can play a major role in early successional forests in a moist, cool region such as Mount Rainier.
Table 19. Successional roles for major tree species on habitat types in Mount Rainier National Park; s = minor seral species, S = major seral species, c = minor climax species, and C = major climax species.
| Habitat type1 | |||||||||||||||
| Species | TSHE/ ACTR | TSHE/ POMU | TSHE/ OPHO | ABAM/ OPHO |
ABAM/ TIUN | ABAM/ VAAL | TSHE/ GASH | ABAM/ GASH | ABAM/ BENE |
ABAM/ XETE | ABAM/ RULA | ABAM/ RHAL | CHNO/ VAOV | ABAM/ MEFE | |
| Abies amabilis | c | c | c | C | C | C | c | C | C | C | C | C | C | C | |
| Abies grandis | S | s | s | s | s | ||||||||||
| Abies lasiocarpa | s | s | S | s | s | ||||||||||
| Abies procera | s | s | S | s | s | s | s | S | s | s | s | s | |||
| Alnus rubra | s | s | s | ||||||||||||
| Chamaecyparis nootkatensis | s | c | c | c | s | c | Sc | SC | C | Sc | |||||
| Picea engelmanii | s | s | s | s | s | s | |||||||||
| Picea sitchensis | s | ||||||||||||||
| Pinus albicaulis | s | ||||||||||||||
| Pinus contorta | s | s | s | ||||||||||||
| Pinus monticola | s | s | s | s | s | s | s | s | s | s | s | ||||
| Pseudotsuga menziesii | S | S | S | s | s | S | S | S | S | S | s | s | s | ||
| Thuja plicata | c | Sc | Sc | Sc | s | Sc | c | c | c | s | s | s | c | ||
| Tsuga heterophylla | C | C | C | Sc | c | Sc | C | c | C | Sc | s | s | c | Sc | |
| Tsuga mertensiana | s | s | Sc | Sc | Sc | Sc | Sc | ||||||||
1TSHE/ACTR = Tsuga heteraphylla/Achlys triphylla, TSHE/POMU = Tsuga heterophylla/Polystichum munitum, TSHE/OPHO = Tsuga heterophylla/Oplopanax horridum, ABAM/OPHO = Abies amabilis/Oplopanax horridum, ABAM/TIUN = Abies amabilis/Tiarella unifoliata, ABAM/VAAL = Abies amabilis/ Vaccinium alaskaense, TSHE/GASH = Tsuga heterophylla/Gaultheria shallon, ABAM/GASH = Abies amabilis/Gaultheria shallon, ABAM/BENE = Abies amabilis/Berberis nervosa, ABAM/XETE = Abies amabilis/Xerophyllum tenax, ABAM/RULA = Abies amabilis/Rubus lasiococcus, ABAM/RHAL = Abies amabilis/Rhododendron albiflorum, CHNO/VAOV = Chamaecyparis nootkatensis/Vaccinium ovalifolium, ABAM/MEFE = Abies amabilis/Menziesia ferruginea. | |||||||||||||||
Pseudotsuga menziesii is the major seral species and Tsuga heterophylla is the major climax species in communities belonging to the Tsuga heterophylla Series at Mount Rainier (Table 19). Abies grandis also plays a minor successional role, as does Alnus rubra on moister habitat types. Interpretation of the role of Thuja plicata is difficult, but we have tentatively assigned it dual roles as a major seral and minor climax species: a major climax role could be argued based on its low mortality rate (see following section) and long life span, factors that should result in increasing stand representation for Thuja over long periods free from major disturbances.
The Abies amabilis Series can be divided into lower and higher elevation segments based on the major Tsuga associate (Table 19).
Abies amabilis is the major climax species throughout. At lower elevations Tsuga heterophylla is a major early successional species and a minor climax species and Pseudotsuga menziesii is a major seral species. Abies procera is a seral species of varying importance depending upon habitat type. Pinus monticola is widespread and can be locally abundant in young and mature stands. Thuja plicata is generally interpreted as a minor climax species on lower elevation Abies amabilis habitats.
Tsuga mertensiana is the major early successional species and a minor climax component on higher and colder Abies amabilis habitats (Table 19). Abies procera, Abies lasiocarpa, Pinus monticola, and Picea engelmannii are typical early successional species, although low-elevation species such as Tsuga heterophylla, Thuja plicata, and Pseudotsuga menziesii may also be represented. The successional role of Chamaecyparis nootkatensis presents problems in interpretation similar to those for Thuja plicata; it is often abundant within stands (hence has a major early successional role), but Chamaecyparis also appears capable of effectively reproducing itself in small numbers and surviving to very old ages.
Few data are available on rates of forest recovery following catastrophic disturbance as compared with our knowledge of patterns of change in forest structure and composition. Rates of recovery are dependent upon many variables, including availability of seed, severity of the post-disturbance environment, and amount of competing vegetation. Recovery can be rapid following small- to medium-sized disturbances on habitats with moderate environments; abundant wind-dispersed seeds are available from surrounding areas of undamaged forest and moisture and temperature conditions are favorable to plant establishment. In such circumstances, reestablishment of forest trees and development of a closed forest canopy can be expected in 20 to 30 years. Competing herb and shrub vegetation can retard forest reestablishment on the most productive (warm and moist) habitats characterized by species such as Athyrium and Oplopanax even in relatively small disturbance areas.
Forest recovery can be especially slow following a large-scale disturbance or a disturbance on environmentally stressful sites. Single wildfires on low- to middle-elevation forest sites often cause rapid regeneration even when they cover large areas; this results from the patchy nature of such wildfires, which often leave individual, groups, and even large patches of streamside corridors of live trees as seed sources (Franklin and Dyrness 1973). Multiple burns of the same area can severely retard forest regeneration, however, by eliminating more and more of the seed source and stimulating competing shrub and herbaceous vegetation. As a consequence, areas subject to multiple wildfires can take many decades or even a century to completely regenerate. For example, Pseudotsuga, Pinus, and Abies procera were still becoming established in the 1886 Cowlitz Burn more than 75 years after the last fire (Hemstrom and Franklin 1982) (Fig. 38). Delayed patterns of regeneration and multiple burns can result in old-growth stands that demonstrate very wide age ranges in the Pseudotsuga dominants (Franklin and Hemstrom 1981).
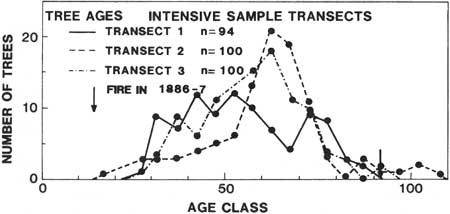
|
| Figure 38. Age-class distribution of trees in stands developed following wildfires in the later 1800's in the Cowlitz River drainage, Mount Rainier National Park, Washington (from Hemstrom, 1979). |
Areas of burned forest at high elevations may also regenerate very slowly, after a single burn and with abundant seed available. Examples can be seen along the Stevens Canyon Road below Mirror Lake and on Crystal Mountain. Such slow regeneration is a consequence of severe site conditions; competing vegetation and animal predation can also be factors in high-elevation burns. Regeneration of some high-elevation sites may need to await a climatic episode that favors tree regeneration, similar to conditions that led to extensive invasion of subalpine meadows early in this century (Franklin et al. 1967).
Tree Mortality and Small-Scale Disturbances
Small-scale disturbances and tree death are important in creating more heterogenous or variable forest conditions. Death of individual or small groups of trees creates critical structures—snags and down logs that fulfill important wildlife and nutrient conservation functions (Harmon et al. 1986, Brown 1985). Mechanical effects, such as the effects of tree uprooting on soil mixing, are also important. Therefore, knowledge of rates and patterns of tree death are valuable in understanding how forest ecosystems function.
Much tree death in the forests of the Pacific Northwest occurs as a result of small-scale disturbances. Such disturbances include small patches or gaps created by windthrow and openings created by contagious diseases, such as the root rot Phellinus weirii, or bark beetles, such as Dendroctonus spp. Mortality of this type may occur continuously or as episodes. Episodes often occur when stands have excess numbers of trees and are subsequently subjected to an environmental stress that renders them more susceptible to beetle or pathogen attack.
Tree mortality has been studied in mature and old-growth forests at Mount Rainier.3 Data were collected from permanent sample plots that have been established in 17 stands representing 13 of the community types described in this monograph; 13 of the plots are in old-growth forest (250 to 1,200 years of age) and 4 are in mature stands (90 to 150 years of age).
3Unpublished manuscript in technical review. "Tree mortality in some mature and old-growth forests in the Cascade Range of Oregon and Washington." by Jerry F. Franklin, Mark Klopsch, Karen J. Luchessa, and Mark E. Harmon. 35 p., 1987. On file at Forestry Sciences Laboratory. Corvallis, Oregon.
Tree mortality averaged 0.52 percent annually over a 10-year period in mature and old-growth forests, indicating a relatively dynamic condition (see footnote 3). Annual rates have fluctuated between 0.27 and 0.86 percent per annum. Major causes of tree mortality are suppression (25.3 percent), unknown (24.6 percent), crushing by falling tree (22.8 percent), windbreak (9.1 percent), and windthrow or uprooting (8.8 per cent). Disease, insects, and miscellaneous causes make up the remainder.
Rates of mortality varied widely with species. Surprisingly, Pseudotsuga, Tsuga heterophylla, and Abies amabilis had comparable rates of mortality (0.57, 0.48, and 0.52 percent per annum). Abies procera had a much higher rate than other common conifers (0.95 percent) for unknown reasons. Thuja plicata and Chamaecyparis nootkatensis had low rates of mortality (0.17 and 0.23 percent per annum). These low rates suggest that Thuja and Chamaecyparis may increase their representation within the forests even though relatively few seedlings and saplings are produced compared to the Tsuga and Abies.
Rates of mortality also varied dramatically with habitat type and with forest age class (see footnote 3). More productive sites tended to have higher rates of mortality. Three old-growth stands on Oplopanax horridum sites averaged 1.08 percent mortality per annum, over twice the average for all plots. Similarly, mature stands (considered to be stands between 100 and 200 years of age) averaged nearly twice the rate for old-growth stands on the same habitat types; such stands are probably still experiencing stand thinning due to competition, the primary cause of mortality in young forest stands.
These data on mortality, along with those from natural forest stands elsewhere in the Pacific Northwest, indicate that tree populations are quite dynamic in mature and old-growth forests such as those at Mount Rainier (see footnote 3). Continuing, modest levels of tree mortality are to be expected, as are occasional episodes with rates up to perhaps 10 times the normal background levels. The basic forest size structure is apparently maintained through such events, and compositional changes occur only very slowly due to low rates of mortality in large, old Pseudotsuga and Thuja, for example.
The gaps or openings created in the forest by tree death and small-scale disturbance at Mount Rainier are too small to allow successful regeneration of the major pioneer species, such as Pseudotsuga. The majority of the mortality occurs as single individuals, and near simultaneous death of more than a half-dozen trees in a small group is very rare. This is a significant ecological feature of the northwestern forests. Small forest gaps are considered to be an important mechanism for perpetuating shade-intolerant tree species in the hardwood forests of eastern North America. At Mount Rainier, the openings are rarely of sufficient size (perhaps 35 m in diameter) to allow reproduction of major shade-intolerant species. Furthermore, forest gaps are typically already occupied by seedlings and saplings of Tsuga, Abies, and Thuja, which utilize the newly available resources. Hence, the shade-intolerant species at Mount Rainier National Park appear to depend upon larger scale disturbances to create suitable conditions for their regeneration.
| <<< Previous | <<< Contents >>> | Next >>> |
chap7.htm
Last Updated: 06-Mar-2007