|
GREAT SMOKY MOUNTAINS
Status of the European Wild Boar Project Management Report No. 6 |

|
PROPOSED RESEARCH
I. Prior Work Outline
Suggested annual data collection (Bratton 1974b).
A. Hogs
1. Yearly census
2. Trapping and shooting records by season
a. Age
b. Sex
c. Condition and weight
d. Number of embryos in females
3. Records of natural mortality
4. Poacher kills
5. Movements into new areas, first records
B. Environment
1. Mast by tree species and elevation
2. Other foods
3. Weather
II. Census
Census of the population will be done by stratifying the population into high, medium, low, and 0 density and then allocating sampling effort accordingly (Cochran 1952; Evans et. al 1966; Goddard 1969). Stratification of the population will be specific to a season. Stratification into different densities will be done through reporting system (visitor reports, ranger reports, hog logs - VPI graduate study) and through a modification of the 5-minute hog sign system for hiking trails (Conley et. al. 1972). Methods for stratification will be standardized. Basis for stratification will be different vegetation types, topographic/moisture and elevation. Potential plots in each stratum will be numbered, and the plots allocated using a table of random numbers. Plots will be 1 km2 in area and shape will be determined by topography. Instantaneous density distribution will be compared with the theoretical Poisson distribution to test for contagion.
Each plot will be sampled by observations and flushing wild boar along transects (line intercept method), following Eberhardt (1968), walked at dusk and after dark with spotlights. This method will only apply to habitats and seasons of high boar densities. Wild boar will be counted and the following information gathered:
A. radial distance from observer to animal,
B. right angle distance from path of observer to the animal,
C. angle of sighting from observer path to point at which animal was first sighted.
The method allows population determination by several estimators, and a Chi-square goodness-of-fit test (Gates et. al. 1968). Gross et. al. (1974) demonstrated methods for determining observer bias in jackrabbit census. Logistical problems may require modification of the random sampling during nights. Data on daily activity patterns may allow evening or even daytime transects during winter. Plots and transects will be permanently located, which will give better estimates of density change than will transects relocated at random each census period (Cochran 1963:283) This system of census should be appropriate to a large park characterized by a great variety of elevations habitats and hog densities. The accuracy of the technique will be unknown but more importantly, precision will be. Precision is more valuable than accuracy when one is interested in trends (Norton-Griffiths 1973).
The method of estimating mobile populations from quick, cursory surveys (Hanson 1968) will be attempted on the boar. The decline in counts as spaces on which animals are successively removed permits estimation of probability of seeing a given animal.
Accuracy of the census will be tested in one relatively small study area in high density stratum through comparison to: 1) capture - recapture methods from trapping (Lincoln Index, modified Schnabel) after Adams (1951) and Ricker (1958), and 2) home range density estimates based upon radio-telemetry. Capture-recapture methods have proven very difficult in the nearby Tellico Game Management Area due to small sample size of returns (Conley et. al. 1972). G. Wood (pers. corres.) has found feral domestic pigs to be alternately trap-shy and trap-prone. In cooperation with G. Wood, A. Tipton (VPI) is presently "working out the bugs" of the capture-recapture method for the South Carolina study area prior to its use in the Great Smoky Mountains.
Indirect census techniques are not proposed. Defecation rates, beds, rubbing trees and rooting are far too variable to warrant any census efforts. These sources of data, however, will provide excellent input for stratum categorization. Counts of tracks in snow during winter will be attempted, if a general snow cover occurs.
III. Radio-telemetry
Using radio-telemetry, population estimates for boar on a small study area can be obtained by delineating the location or boundaries of discrete groups of boar and then determining the number of boar per group, similar to density estimates for mountain lions in Idaho (Hornocker 1970) and timber wolves in northeastern Minnesota (Van Ballenberghe et. al 1975). The amount of group interchange and the proportion of single boar will need to be known. This grouping in formation can be obtained through: 1) census in formation, 2) the trapping program and 3) sight locations of radioed boar within their group(s).
Number of radioed boar should be sufficiently large and should incorporate the home ranges of more than one discrete group (for comparisons). Usually time and costs limit the number of radioed animals 13 or less. Dr. G. Wood (Baruch Institute, Clemson University) suggests maintaining 5-7 active radios on feral domestic pigs, and cautions that some radios are lost through malfunctions and self removal (pers. corres.). Radio-telemetry should also provide precise habitat use information in a small area for comparison to habitat relations determined through census. The quantity of particular vegetation or habitat types can be determined within the home range of a radioed boar and the time spent within each type determined by repeated locations. Preference or avoidance of types can be determined by a series of Chi-square tests comparing availability with utilization (Neu et. al. 1974).
Monitoring devices within the radio collar (AVM, Champagne, Illinois), will provide information on daily activity patterns. AVM radios, receivers, and monitoring devices have been recommended to me by G. Wood, M. Hornocker, Canadian Wildlife Service and Alberta Parks, Recreation and Wildlife.
The study area for census and radio-telemetry should initially be excluded from all control activities. Once baseline data is established, intensive hunting and trapping in the area will provide essential information on reaction of boar to control effort through; 1) movements, 2) changes in daily activity patterns, and 3) changes in habitat and cover use. The control effort should also bring in a large sample of marked animals.
Radio-telemetry will also provide specific needed information on seasonal movements. Belden (1972) has already described a general movement of boar to higher elevations during the summer months followed by reverse movement in the fall. Information is needed on the causal relationships for these movements, the exact direction and locations of boar leaving the high elevation beech gaps where damage is most severe, and the magnitude of movements. This information should help to focus winter control work on key areas. The study area and season should be designed specifically to gather this information.
IV. Eruption Stages
Eruptive fluctuations of populations initiated by a liberation have been described in New Zealand by Riney (1964), Howard (1965) and Caughley (1970). The four hypothetical stages, defined only by attributes of the population are:
A. initial increase
B. initial stabilization
C. decline
D. post decline
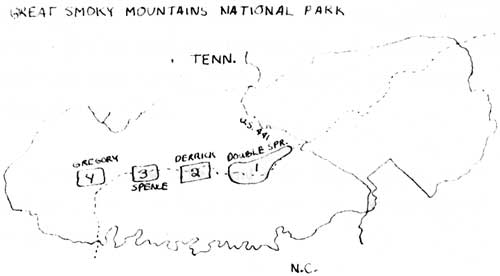
The sequence following liberation can be observed both at one point over a range of time, and at one time over a range of distance. The model predicts that stage 2, the "initial stabilization" stage, will be found about midway between the dispersal front and the area containing the stage 4 population. This model will be tested for the wild boar in the Great Smokies through intensive sampling of boar in one vegetation type, the gray beech forest along the central upland of the park. Stages 4 and 1 were identified from the initial liberation point and recent invasion, respectively; stages 2 and 3 were chosen on their relationship to the other two stages (Figure 5).
|
| Figure 5. Hypothesized eruptive stages for European wild boar within gray beech forest, GSMNP. |
The gray beech forest is the vegetation type with greatest hog damage and will be a focus of control efforts. The large number of animals expected to come from these areas will be investigated for differences in fecundity, survival and fat reserves. By hypothesis, the rate of increase at Indian Gap-Double Springs sample should be positive, rates at Derrick and Gregory should be close to zero, and the Spence Field rate should be negative (Caughley 1970). Fecundity, survival and fat reserve statistics should be highest at Double Springs-Indian Gap, lowest at Spence Field, and intermediate for Gregory and Derrick. The model also predicts a decrease in food supply from Double Springs to Spence Field; this hypothesis will be tested by sampling plots for: 1) declines in preferred forage for the boar along the line and, 2) increases in plants not eaten or ones that thrive in the disturbed areas. Extension food habits information will be required from the four sample areas.
The strong influence of weather and mast crop on boar reproduction will presumably be held constant for samples taken along this line within the same year.
The eruptive states for boar in the (Great Smoky Mountains may be difficult or impossible to identify. Klein (1968) suggested that ecosystem complexity is related to eruptions; an environment with several diverse population regulating mechanisms would bring about a more gradual control of a species. Estimation of population parameters is difficult in the boar because of: 1) high variance for season of births, 2) high frequency of multiple births, 3) possible confusion of pattern of spread, and 4) complex, omnivorous food habits. The region east of Highway 441 was excluded because of the possibility of several points of liberation.
The exercise of attempting to identify the eruptive stages will: increase knowledge of the population movement, help to relate control effects to natural rates of increase, and help to precisitate other strong influences on population parameters.
V. Behavior
Wild pigs are more difficult to study than other ungulates due to adoption of nocturnal habits, intelligence, shyness, alertness, and acute senses of smell and hearing (Frädrich 1974). Of the eight species of wild pigs, only the behavior of the European wild boar and the African wart-hog (Phacocherus aethiopicus) have been carefully studied in the wild, and even in the case of these species information is inadequate.
Specific behavioral studies on European wild boar have been conducted by Mohr (1960) and Porzig (1966). African wart-hog behavior has been researched by Geigy (1955) and Frädrich (1965). A comparison of behavior in the Suidae is provided by Frädrich (1975). Many aspects of social behavior in the European wild boar, including group sizes, mating behavior, and farrowing behavior can be found in the more comprehensive works on boar (Boback 1957; Sludskii 1956; Niethammer 1963; Heptner and Naumov 1966; and Snethlage 1967).
Basic information is needed on daily activity patterns of boar, on group size of boar, seasonal changes in group size, familial relations in groups, average frequency of association for different sex and age classes in groups (group cohesion), dominance relations, leadership of groups, and extensive data on breeding behavior. In particular, data is needed on the breeding behavior and strategies within groups and the relative input of certain males to the total conception rates. This information will provide insight into more effective direct control and indirect control measures (poisons, sterilization, birth disrupters) and the best times and methods for transmission.
Certain largest and strongest boars are suspected of siring most piglets (Stegeman 1968; 286). Kurz and Marchinton (1972) reported that males in breeding groups of feral domestics established dominance hierarchies; on one occasion, a larger male drove away a small male that was attempting to breed a receptive sow. In domestic swine, males were dominant to sows, and dominance within sexes was significantly correlated to an animal's weight (Beilharz and Cox 1967). Frädrich (1974) reported that European wild boar (males) usually cannot mate until they are 4 to 5 years old due to competition with other males, even though they reach sexual maturity at 8 to 10 months.
Information is needed on dominance in relation to the first animals to enter traps, take poisons, and the effect of selective removals, if any, upon reproductive success. Selective removal of mature bull elk (Cervus canadensis), for example, has had no effect upon the Yakimaw (Washington) elk herd; pregnancy rates and productivity of elk remain the highest recorded in North America even though almost all the breeding is done by a very small number of yearling (spike) bulls.
Further research on the social behavior of wild boar might be questioned, in view of the amount of information already available. Justification for the research includes: 1) the social behavior information available is not specific enough to allow direction of the control program in the Park, 2) information is needed on the presence, location and timing of any large boar concentrations in the Park that might be group-trapped, 3) the reviously mentioned information on breeding stategies is needed to direct any specific chemicals and, 4) the specific social behavior, population regulation and feral domestic vs. wild boar relations in the Great Smoky Mountains National Park should be known.
It is hypothesized that the European wild boar possess greater population regulatory mechanisms than domestic swine, and, therefore achieve lower densities and cause less vegetation damage. Comparisons of these features between the two types are needed. The population in the Great Smoky Mountains is known to contain European wild boar, feral domestics, and hybrids, based upon coloration patterns (Fox 1972; M. Shaffer pers. corres.) and this statement is supported by chromosome work in nearby Tellico (GMA (Conley et. al. 1972). This information is needed should it be desirable to shift the population to wild boar-type or feral domestic-type; this shift is obviously possible since 31+ years of "natural" selection has not eliminated the genotype of either. European wild boar-type will undoubtedly be selected for under severe hunting and trapping pressure, especially with hunting dogs, and under any significant timber wolf predation. Selection for the wild boar-type will be accelerated by elimination of feral domest ic sources along park boundaries. The feral swine-type can be encouraged through actual transplants within the park and the continued lack of natural predators. The European wild boar-type might occur at lesser densities, and do less damage, but the feral domestic-type might move less and might be easier to trap and shoot.
Behavior work should follow two steps or phases: 1) ongoing, preliminary phase, 2) a later intensive study using marked animals. The first phase can be conducted during census of the park and will provide general information. Specific information on group cohesion, dominance relations and leadership will require a separate intensive study involving a large sample of marked animals and observations following Hawkins and Klimstra (1970). This second intensive study might well be integrated with the radio-telemetry study, in order to repeatedly approach radioed boar to investigate their group relations. This method would solve the problems which were encountered in the summer of 1976 trying to repeatedly observe animals in dense tree/shrub structure—without the added bias of baiting pigs to a traditional site. No one has yet done a behavior study of boar using marked animals.
VI. Competition
Matschke (1965) reported that wild turkey (Meleagris gallapavo) and ruffed grouse (Bonasa umbellus) maintain numbers in spite of introduction of European wild boar into an area. Predation of nests was the same on high and low hog population areas, but in the high population area, the hog destroyed the most nests. Henry (1968) reported that the wild boar is a very minor predator (of dummy nests) and does not add additional predation to nests, but replaces predation by other species. In the justification section, I mentioned speculations on the 1) overexploitation of salamander and reptiles by boar, 2) avoidance of boar by white-tailed deer and 3) dietary competition of boar with other wildlife.
The effect of wild boar on terrestrial animals with small home ranges can be tested with two study plots and the erection of an exclosure on one. Both plots should be sampled prior to exclosure being erected, and then in succeeding years. In that way the control plot will serve to correct for responses of animal populations to factors other than wild boar. Exclosures should allow free movement of all species other than boar. Plots and exclosure should be specific to certain species and the location determined by the species. Rocky groundwater seepage and streambeds with a heavy density of wild boar could be used for a salamander check; rock sections in the drier North Carolina side of the park for a rattlesnake test; etc. Consultation with specialists in herpetology will be required and services of a volunteer or student may be solicited for 1977.
A study of competition through food habits alone is precluded by wild boar rooting since it is destructive or effective to many other plants than the one being eaten. The degree to which hog feeding sites are or were sites to be potentially used by other wildlife (black bear, white-tailed deer, wild turkey, ruffed grouse, and gray squirrel) is best evaluated by a description of the distributions, habitat relations, and interspecific associations of all species. In cases of a significant habitat overlap, further research on food habits and specific effects of the rooting can be conducted in those areas, Interspecific associations can be evaluated by several methods (Cole 1949).
Lack of overlap in habitat use and food habits does not preclude the existence of competition at some time in the past, which has since been compensated for. This possibility can be investigated by establishing distribution, population size and habitat relations of species in the southeastern quarter of the park prior to boar invasion. Significant habitat and dietary overlaps are not generally considered competition unless a decline occurs in one species, or the co-action occurs on a forage in limiting supply (Hutchinson 1965).
Any data on competition between the exotic boar and other species will add perspective to the management problem, and will pinpoint sensitive locations and habitats for control work.
VII. Habitat Relationships
Belden (1972) described rooting and wallowing changes of boar through a 1-year period in the Great Smoky Mountains National park and adjacent Tellico GMA. Altitudinal shifts occurred during summer, to higher elevations, with a return to lower elevations in winter. Rooting was most common in cove hardwood in spring, northern hardwood in spring and summer, and closed-oak type during fall. Vegetational occurrence and elevational movement were related.
The attached data sheet (Appendix) has been prepared for observations of wild boar taken during systematic portions of the census work. Information recorded should allow habitat relation of boar through both classification and gradient schemes for habitat. The surveys should have the following features:
A. Systematic surveys where all boar and other wildlife are consistently recorded. Vegetation types, elevations and topographic/moisture features should be sampled in proportion to their occurrence on the study area(s).
B. Acreages of vegetation types, topo/moisture features etc. must be known, since percent use means little without a comparison to availability (Neu et. al. 1974).
C. Boar must be equally observable in all types, or a correction factor used for the more open types, if this assumption is found false.
D. If boar sign is recorded, a correction factor should be employed for greater observability of rooting vs. tracks vs. grazing.
E. Sample sizes will necessarily need to be large to detect differences between years and to detect preference or avoidance of different vegetation types. Preliminary results may be crude. Information on major forage species will be gathered after habitat and dietary relations are established. Ultimately a model of boar habitat along environmental gradients will be constructed, the niche of Hutchinson (1965), following example of Singer and Ackerman (in progress).
VIII. Seasonal Food Habits/Forage Relations
The niche or functional role of the European wild boar in Great Smoky Mountains National Park is poorly understood. More data is needed on the interaction of wild boar with major forage species. Certain rizomatous or bulb-reproducing plants may be sustaining themselves, while other perennial and seed-producing plants may be declining in heavily rooted sites. The growth rates and productivity of specific forage plants should be described. The successional relationships of boar can be quantified by aging the feeding site - i.e., old homesite, pasture, logged or burned stage, virgin forest. The successional status of the forage plants should be described. The interaction of wild boar with major forage species can be partly estimated during the sampling for eruptive stages.
It is hypothesized that since the wild boar has a short digestive system it functions in an energy rich or protein niche, using food sources high in stored fats, carbohydrate, protein, or sugars; for example, animal matte , invertebrates, tubers, roots, acorns, fruits, berries, and forbs from fluesic sites. The interspersion of these types of items in an area undoubtedly influences boar densities.
The effect of boar on the timing and sequence of energy flow in forest ecosystem is unknown. This can be determined by data on: 1) forage standing crop, 2) net production of forage, 3) ungulate standing crop and secondary production, 4) energy flow through ungulate population, and 5) comparisons to an equivalent area without a hog population.
Food habits will be studied through stomachs collected during the control program. In addition, an effort will be made to collect a minimum of two specimens in each habitat with major use during each of four seasons in a 1-year period (October 1976 - October 1977). This will require research sampling additional to control work. Low elevation food habits need not be stressed in the sampling due to Scott's (1974) work. Additional food habits information will be gathered, where appropriate by the feeding site method (Cole 1956; Knowlton 1960). Percentages for individual forages species will be aggregated and averaged for all sites (Martin et. al. 1946); this method will only be appropriate to the boar and study area during periods of snow or appropriate ground cover. Where animals are readily observed, the time-observation method (Buechner 1950; Harper et. al. 1967) will be employed. Visual estimates of the utilized vegetation will be made after the boar vacates the site.
The combination of three methods is viewed as the most appropriate manner to study food habits. Stomach analysis alone is limited for species identifications and a bias towards larger and less digestible food items.
Macro-identification of stomach contents will follow flushing through a four-member gang sieve with mesh sizes of 1, 2, 4 and 8 mm (Cushwa et. al. 1970). Dry weights of separates will be determined at 70° C. Histological techniques and a reference slide collection will follow Baumgartner and Martin (1939). General approach follows that used by G. Wood at Hobcaw Colony, South Carolina, the results of which are expected soon in a Master's thesis.
Documentation of plant species dominance and distribu ional chances in relation to the boar invasion will provide an opportunity for a study in natural selection among plants.
IX. Management of Natural Predators
"Insofar as possible, control through natural predation will be encouraged" (U.S. Dept. Interior 1970).
Within its native range, the European wild boar has evolved since pre-Pleistocene times in the face of significant predation from brown bear (Ursus arctos), wolf (Canis lupus), tiger (Panthera tigris), and leopard (Panthera pardus), often in conjunction with human predation (Bratton 1974b , citing Rakov l970). Sloudskii (1956) estimated that an adult wolf can kill 50-80 boar (mostly young) in a year; boar can comprise as much as 22-35 percent of the wolf diet. Boar are particularly vulnerable to wolves in deep snow, since the wolf weight load is 160 g per square cm, while the boar load is 251-478 g per square cm. In order, wolves kill young first, then yearlings and then adult females; adult males are difficult prey and actually can kill wolves (Sludskii 1956). Adult females attempt to valiantly guard young during their first 10 months and often fall prey to wolves while doing so.
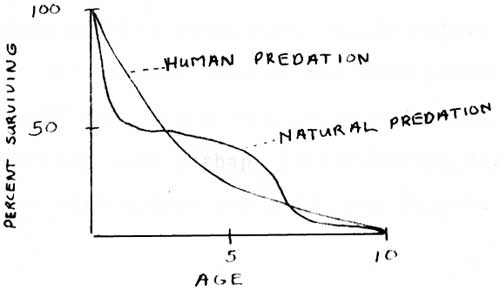
A comparison of human predation and large carnivore predation upon a large herbivore is provided in Figure 6, after Krantz (1970) The carnivore data is based on studies of wolves (Murie 1944; Mech 1966). The human predation curve is approximated by the control program in the Great Smoky Mountains National Park (see Figure 4). in addition, total of 259 wild pigs (Sus scrota) killed by hunting in Perak approximated the age class ratios of the wild pig population (Diong 1973).
|
| Figure 6. Comparison of survivorship of a large herbivore under human predation and natural predation (Krantz 1970). |
The curves demonstrate that the number of surviving young adults, the major breeding stock, is more numerous under human predation because their numbers are not so drastically depleted when they are still immature. When natural predators are replaced by human hunting the most likely expectation is an increase in the prey population.
The subject of a wolf reintroduction into the Great Smoky Mountains is a matter of discussion. Information is not available on the prey base present to support a wolf population. Even if space and food requirements are present the region is populated, with major human centers occuring on the Park boundary at Bryson City, Cherokee and Gatlinburg. Public relations may become a serious problem since many people naturally dislike wolves especially if they have to tolerate problems associated with a dense wolf population such as depredations upon domestic livestock including (and perhaps particularly) dogs and cats, and competition with wolves for wild game (Van Ballenberghe et. al 1975).
Effects of the wolves on prey species needs to be predicted. The function of the system after a major perturbation, such as a hog decline after a poor mast year or hog eradication, should be predicted. Lynx (Lynx canadensis) in Newfoundland, greatly increased and were able to exert a detrimental effect upon caribou (Rangifer tarandus caribou) through excessive calf predation, after introduction of a new prey species, snowshoe hare (Lepus americanus) (Bergerud 1971).
The size of Great Smoky Mountains National Park, adjacent National Forest lands and the presence of white-tailed deer and European wild boar justify an investigation of potential timber wolfe reintroduction. Van Ballenberghe (1975) found pack territories of only 52 to 145 km2 (one wolfe per 23.6 km2), and rendevouz sites of two adjacent packs only 1.6 km apart in an area of high wolfe density in Minnesota. Pimlott (1967) suggested 3.9 deer per km2 with an annual productivity of 37% would be required to support a population of one wolfe per 25.9 km2 (in Algonquin Park, Ontario). The goal for the wolfe population in Minnesota is one wolfe per 10 square miles, which requires a spring ratio of 80 deer per wolfe or 8 deer per square mile (Eastern Timber Wolfe Recovery Team, in press).
The Eastern Timber Wolf Recovery Team identified the following areas for wolf re-establishment possibilities: 1) northeastern Maine, 2) northwestern Maine, 3) White Mountain area, 4) Adirondack Forest Preserve Area, 5) Southern Appalachians (northern section), 6) Southern Appalachians (southern section), 7) Upper Peninsula of Michigan and 8) northern Wisconsin. The team identified work to be done in each of those areas to determine whether re-establishment is socially and ecologically sound:
1. Consult vegetation and ownership maps, land use maps and plans, and local biologists to define and select all suitable areas for transplant.
2. Determine deer, moose and beaver density in the selected areas (in our case boar).
3. Determine human densities and use patterns in the selected areas.
4. Determine possible impact of transplant on public health.
5. Estimate effect of establishing wolves on other wildlife and domestic animals.
6. Determine legal implications of transplant.
This work is programmed and will be completed by January 1977 along with a compilation of all mountain lion (Felis concolor) sightings in the area, and evaluation of any trends or key areas in the lion sightings. The eastern mountain lion is listed as an endangered species under the Endangered Species Act of 1973 and its critical habitat and continued existence matters of law for all Federal agencies.
Determination of prey densities, prey distributions, prey vulnerability patterns, human density patterns, and livestock distributions is required for both mountain lion perspective and timber wolfe potential.
X. Control
Poisoning and Sterilization. The comments of Bratton (1974b) are still appropriate; both poisoning and sterilization will require extremely specific baits. This will require a more extensive natural food habits and artificial food preference study.
Season of Control Work. In adjacent Tellico GMA, farrowing peaks for European wild boar occur in mid-winter (January and February) and early summer (May and June) (Henry 1966). Natality is highest during December - March and litters are larger. However, most hogs killed during hunts are born April to July, which suggests higher mortality among winter-born hogs. Therefore, the April to July period has the lowest number of juveniles per female and most sows will be pregnant or suckling. Any emphasis in control work should occur April to July or prior to April.
Control work should be intensified during the winter andi spring following fall of poor mast production, since few or no sows will farrow at this time (Matschke 1964). Ongoing research contracted to Virginia Polytech Institute (VPI), will attempt to identify mast crop fluctuations.
Trapping. Trapping of wild boar is successful in Russia to control populations when done once a year with large corral traps (B. Evison pers. corres.). Winter conditions vary between the Russian and park area which may effect concentrations of boar and the attraction of bait as food. R. Conley (pers. corres.) reported that wild boar remain dispersed throughout the winter at Tellico GMA and would not be attracted to a permanent trap. The method merits an attempt in the Great Smoky Mountains where it may prove successful during poor mast years and/or severe winters. Wire noose snares in Perak are successful to the point of jeopardizing wild pig populations (Diong 1973), but their non-selectivity precludes their use in the park.
Trap success rate has been low in the park, ranging from .0062 - .0328 captures per trap night (Duncan 1972). Higher rates were obtained when recent hog activity was located near the trap site, and after hogs had begun to accept bait. A new trap based on the "Telgazlie" trap (Fraser 1975) was built at the Uplands Field Lab in April 1976 and has shown initial promise of higher success. Multiple captures have not yet been made and this should be a primary objective (Bratton 1974b). The new trap is much easier to transport into the back country and the open top allows other wildlife (bear, turkey, deer) to escape without injury. A review of trap designs is provided in the Appendix.
Shooting. Reduction by hunting is successful in controlling wild boar populations in Europe in many areas (Henning 1963, cited in Bratton 1974b). Sport hunting can place heavy pressure on wild boar in nearby Tellico GMA in years after a poor mast crop (R. Conley pers. corres.).
Heavy hunting pressure is harder to obtain with government shooting operations because of the cost of fielding equivalent forces. Anderson and Henderson (1961) state; "Investigations generally into government shooting operations have led to the conclusion that undesirable population surges have been induced in many of New Zealand's wild animal herds as a direct result of the application of spasmodic and fluctuating shooting pressure." Howard (1964) stated: "There are a number of un proven instances that illustrate how the 'predation' resulting from a control program of government hunters, shooting only where and as long as large tallies could be reported, resulted in stimulating an even greater density of deer, with consequent additional damage to the habitat (in New Zealand)." Unless cropping of elephants is by complete family groups only, larger groups result and vegetation damage increases (Laws and Parker 1968).
Dog Hunting. Hunting with dogs may simulate natural predation in that intensive hunting results in greatest losses of young, then yearlings, then sows, then boars (Sludski 1956). Herds can be dispersed with hunting in large groups of 20 to 30 dogs; boar moved 15-20 km from a region after 2 days of pursuit. Many problems would result from dog use in the park with control of the dogs, chasing of other game, public relations and stimulation of poaching interest. Dogs might find limited use, however, in the severely damaged beech gaps in spring when they would be disruptive to farrowing and young piglets would be vulnerable. Sweeney et. al. (1971) found that white-tailed deer chased by dogs left their home ranges in 5l of 65 chases, but in all but one case, returned within 1 day or less.
Compensatory Mechanisms. The population of European wild boar in the Great Smoky Mountains is likely to respond to the control effort. Changes may occur in the age of breeding, number of young produced per sow, survival of young, daily activity patterns, movements and habitat use. During an intensive shooting control program on duiker in Africa, the population responded through greater breeding and more juveniles breeding (the number of calves doubled), and altered their daily activity pattern toward evenings (Wilson and Roth 1967). There were no significant effects upon population size, movements, or the age structure, in spite of the fact that the hunter density was one per 5 square miles for the entire 2-year period. Fear is a response which is easily transmitted and retained in ungulates. Response of fleeing continued for a long time after hunting stopped in red deer (Batcheler 1968). The hunted duiker retained their altered crepuscular pattern for 1 year after control shooting stopped (Wilson and Roth 1967).
Behavioral compensation can be to the advantage of the control program. Chamois in New Zealand altered their habitat use to less optimal, rugged terrain under severe shooting pressure, and this greatly reduced their reproductive compensatory mechanisms (Butcheler 1968). Unfortuantely, Geist (1971) predicted that loss of favorable habitat due to disturbances will likely be insignificant in species characterized by high reproductive rates, short life expectancy and the ability to adopt nocturnal habits (i.e. wild boar).
The reproductive capability of wild boar is considerable. Annual increases of 178 percent have been reported in the Belowesh Preserve (Kozlo 1970). Higher fertility and fecundity of adult doe and a higher proportion of pregnant fawn white-tails was found in an area of New York where population density was lower and forage production higher (Cheatum and Severinghaus 1950). Reduction of the elk herd in a watershed in the Blue Mountains of Washington approximately doubled the amount of available forage per individual and resulted in a 50 percent pregnancy rate among yearling cow elk (Buechner and Swanson 1956).
Documentation of the response of the boar population to control can be conducted along with the control effort. Baseline data on daily activity patterns, habitat use, group behavior should be gathered prior to and after intensive control work in the area. Radio-telemetry will provide information on movements, changes in daily activity patterns of individual boar, and changes in habitat and cover use of individual boar in response to control efforts. Population response can be monitored throughout the control effort.
XI. Effect of Wild Boar on Water Quality
European wild boar regularly wallow in small streams, springs and stream head waters. Boar turn over rocks and root up small stream beds, presumably searching for invertebrates and salamanders. Boar also root along the banks of larger streams and rivers. The preference by boar for mesic sites is well documented in the park, including preference for more mesic sites in Gray Beech Forest (Bratton 1975), and the low-lying, more mesic sites in the agricultural Cades Cove fields (Fox 1972). This preference repeatedly brings the boar closer to water-courses.
The full or part-time services of a water quality specialist are soon expected at the Uplands Field Lab. This person would be better qualified to evaluate the effects of wild boar on stream quality. In the meantime, the Principal Investigator will continue to take field notes on locations and extent of drainage rooting by boar. Preliminary work on algal communities in rooted and undistrubed watersheds has been awarded to Dr. Rex Lowe, Bowling Green University, 1976-78.
XII. National Seashores in the Southeast Region
Gulf Islands National Seashore, Mississippi, and Cumberland Island National Seashore, Georgia, contain populations of feral domestic hogs. Hogs were introduced in 1949 to Gulf Islands (specifically Horn Island) to provide a self-sustaining source of food for personnel at the military base there. Feral hogs on Cumberland Island are escaped domestics; other livestock include cattle and horses.
Feral hogs feed heavily on roots of many plants, including
N1 = R2 - RP2 P1 - P2 where, Xi = number of x-type animals in pop. at time ti
Yi = number of t-type animals in population at time ti
Ni = Xi + Yi (total population size at time ti)
Rx = X1 - X2
R = Rx + Ry (=N1 - N2)
Pi = Xi/Ni (= 1 - Qi)
Estimates from Census. Transects walked by the line intercept method (Eberhardt 1968), population stratification, stratification by recognizable vegetation types and other methods being developed in the Great Smoky Mountains National Park. Aerial census might prove appropriate, especially if the hogs regularly use beach areas in numbers. Crocodiles basking along river banks were censused from aerial photos taken from a helicopter (Parker and Watson 1970); use of reference stakes allowed size/age determination and construction of a survivorship curve. Detection was made easy in high marsh grass when the animals spooked in response to the low-flying aircraft.
Knowledge of the population, sex and age distribution and control data will allow estimation of population size following Wood et. al. (in press) as used in the Pennsylvania deer harvest.
Estimate from Trap/Retrap. Population estimation through trap/retrap efforts on feral domestic hogs is being conducted by G. Wood at the Belle Baruch Forest Science Institute. Dr. Alan Tipton (V.P.I.) is helping to analyze the data as part of a cooperative effort with Tipton's eventual purpose of applying the data to the Great Smoky Mountains National Park. This type of co-operation should be very strongly supported.
Application of the knowledge gained at Baruch will be applied to the Smokies through Tipton's contact there, and hopefully, through acquiring consultation services of Dr. Wood on Cumberland. Cumberland and Horn Islands are similar situations to some extent. The techniques developed at one island can be applied to the other. Both islands are closed populations and provide ideal study situations to test and verify census techniques being used in the Smokies. Horn Island (hogs) and Petis Bois (no hogs) provide a classic controlled situation to study effects of feral hogs on vegetation and animals.
Gathering of data in the three study areas—a small island, a large island, and a diverse, relatively stable forest ecosystem—will provide insight into the density, vegetation damage, and rates of increase in wild hogs for situations varying widely in: 1) size and dispersal rates, 2) successional status, and 3) primary productivity.
Central co-ordination at the Uplands Laboratory in Gatlinburg will maximize expenditures and efforts in all three units. An effort will be made to maximize the speed, efficiency and cost of the research effort through organized use of the staff Regional expertise (Dr. S. Bratton — plant ecology; Dr. D. Stoneburner —populations; F. Singer — wildlife) and any additional staffing. Hired consultants will be employed if possible in situations where it is an obvious time and financial saving.
| <<< Previous | <<< Contents>>> | Next >>> |
res/6/sec6.htm
Last Updated: 24-Aug-2009