|
CUMBERLAND ISLAND
An Ecological Survey of the Coastal Region of Georgia NPS Scientific Monograph No. 3 |

|
CHAPTER 5:
The Open Marine and Estuarine Waters
The marshes, being protected by the barrier islands from frontal assault by the ocean, interact with the ocean indirectly, through systems of tidal streams and estuaries (Fig. 26). Estuaries are semi-enclosed bodies of water having free connections with the open sea and having sea water measurably diluted with fresh water derived from land drainage (Pritchard 1967a). The estuaries of Georgia connect with the sea through the sounds that separate the barrier islands. These are north to south from the Savannah River Entrance to the St. Marys River Entrance: Wassaw Sound, Ossabaw Sound, St. Catherines Sound, Sapelo Sound, Doboy Sound, Altamaha Sound, St. Simons Sound, and St. Andrews Sound. Salt water is diluted by fresh water from the Savannah, Ogeechee, Altamaha, Satilla, and St. Marys rivers. The coastal or inshore waters extend from the mouths of the sounds and from the barrier beaches to a depth of about 60 ft. The offshore waters of the continental shelf lie beyond.
The continental shelf along the Georgia coast is 70-80 miles wide with a gentle slope of about 2 ft per mile (Henry and Hoyt 1968). It is covered by sediments deposited during low stands of the sea. The topography of the continental shelf off the Georgia coast is shown in Fig. 29. The poorly defined valley in the shelf east of Sapelo Island is thought to be a filled valley once occupied by the Altamaha River (Pilkey and Giles 1965). An abrupt increase in slope of descent occurs at 50-80 m (Pilkey and Giles 1965). This break marks the edge of the continental shelf and the beginning of the upper continental slope. Seaward of the continental shelf is the Blake Plateau, an intermediate plateau between the continental shelf and the ocean basin. The Blake Plateau averages 700-1000 m depth (Pilkey and Terlecky 1966).
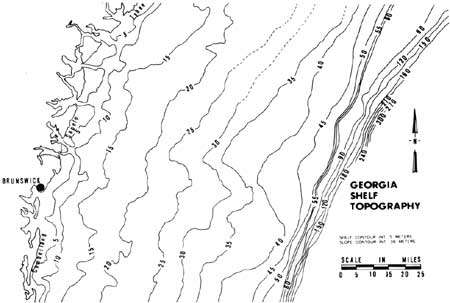
|
| Fig. 29. Bottom topography of the continental shelf off Georgia. Contour interval changes at continental shelf. (Adapted and redrawn from Pilkey and Giles 1965.) |
Physical characteristics
The coastal (inshore) waters are somewhat diluted with fresh water; they are turbid and most productive. Beyond the continental slope, ocean waters are clear and relatively unproductive. The waters of the continental shelf are of intermediate fertility.
Patterns of circulation
The waters of the Georgia coast are subject to many interacting forces that produce complicated patterns of circulation that to a large extent determine the distribution of sediments, nutrients, oxygen, temperature, salinity, food, and planktonic forms of larval and adult organisms.
Ocean currents are produced by wind, gravity, and differences in density of water strata. Surface currents commonly are caused by the stress that winds exert on the water surface; tidal currents are caused by the interacting forces of gravity of the earth, the moon, and the sun; and density currents are related to differences in density that result from variations in temperature and salinity within a body of water.
The Gulf Stream, a northerly flowing current of warm, saline water beyond the continental slope, is a density current. It results from warm, tropical waters spreading northward over the more dense polar waters which sink and flow toward the equator. The currents are deflected from direct poleward flow by wind friction, earth rotation, and land masses. Eddies form inshore from the Gulf Stream and may be responsible, in part, for the southward transport of sediment along the Georgia coast.
The movement of the waters inshore from the Gulf Stream has not been studied systematically and must be assumed from a limited number of observations. Bumpus and Lauzier (1965) summarized data from drift bottles on seasonal direction of surface currents of the continental shelf from Newfoundland to Florida. The data from Georgia, admittedly inadequate, indicate currents flowing as follows: winter—southerly; spring—northerly; summer—southerly and northerly; and fall—southerly. Bellinger (1968) reports that flow is southward during the summer, fall, and winter, extending out 20-30 miles during the summer and about 45 miles during the other seasons. Rates of surface flow vary from 2 to 10 miles per day; bottom currents flow at about 1/10 the rate of surface currents. Superimposed on the southward circulation is either an inshore surface current and an offshore bottom current or vice-versa. Bellinger (1968) reports that there are no data on the current flows between the Gulf Stream and the longshore current.
The surface currents are mostly dependent on the direction of the wind. The wind is predominantly from the northeast and northwest in the winter (December—February), from the south in spring and summer (March—August), and from the northeast in the fail (September—November) (Carter 1959; U. S. Naval Oceanographic Office 1963; Kuroda and Marland 1969). These prevailing winds coincide with the direction of the surface currents, and seasonal differences in wind velocities are partly responsible for the longshore movement of sediments southward. The northerly winds of the winter and fall are much stronger than those of the summer and spring (Neiheisel 1965).
As previously stated, the Georgia coastline is reported to have the lowest energy levels along the southeastern coast (Tanner 1960). Neiheisel (1965) confirmed this by analysis of wave refraction diagrams. The shoreline is aligned at about N23°E; the continental shelf is broad; the surf is mainly from the southeast, and when waves approach shore from this direction they are refracted, resulting in an approach parallel to the shore. During this process, energy is dissipated over a broad area. Waves approaching from the northeast, the direction from which the strongest winds and the highest seas approach, are refracted more and energy reduction is greater. Neiheisel (1965) concludes that the low energies along the Georgia coast are due to the wide continental shelf and the gentle offshore slope which cause incoming waves to be refracted more, and to the fact that the greatest refraction of waves occurs when wind is from the direction of strongest force.
Density differences caused by seasonal changes in salinity and temperature also produce water movements. Depth profiles made during cruises of the R/V Gill, the results of which have been reported by Anderson and Gehringer (1957, 1958, 1959) and Anderson et al. (1956), provide the basis for the following discussion. Inshore waters have temperatures of 29°C and salinities of 35 ppt in August and are less dense than those at the shelf break which have similar temperatures, but higher salinities. By November, inshore waters are cool, while the offshore waters remain warm because of the influence of the Gulf Stream. High salinities prevail throughout, reflecting the low fresh water input from June through September (U.S. Geological Survey 1960). Inshore waters are now more dense than offshore waters. In February waters are colder both inshore and offshore; inshore waters are less saline because of greater fresh-water runoff; and the difference in the densities of the water masses is less. As temperatures begin to rise in the spring, inshore waters warm faster and the temperature gradient gradually disappears, but the salinity gradient is steeper than at any other time of the year because of heavy freshwater discharge. The lessening temperature gradient and steepening salinity gradient compensate to produce water of similar densities inshore and offshore. These shifts in densities probably result in slow, seasonal water movements. They could be responsible for setting up the inshore-offshore components mentioned by Bellinger (1968).
Tidal currents are the horizontal movements of water which, unlike the currents caused by wind and density differences, are periodic. The tidal component is most pronounced in narrows such as entrances to bays and constricted parts of rivers. Tidal currents for the Georgia estuaries were given by Haight (1938).
The Georgia coast is subject twice daily to tides approximately the same height. The height of the tide increases from about 5.5 ft at Nassau Sound, Fla., to about 7 ft near Savannah. The tidal movement of saline waters into the estuaries and the drainage of rivers into them cause the complicated hydrologic patterns characteristic of estuaries.
Pritchard (1967b) described four hypothetical circulation patterns for estuaries, two of which may be applicable to Georgia estuaries. The moderately stratified estuary is one in which tidal action serves as the dominant force mixing fresh and salt waters. The Savannah River exhibits this type of circulation. In an estuary with no tide or friction, undiluted sea water would extend upstream along the bottom to a point where the river surface was approximately at sea level. The less dense fresh water would flow seaward on top of the salt water. When there is tidal action, as in the moderately stratified estuary, turbulence carries fresh water downward and salt water upward. The salt content of both layers increases toward the sea, but at any given point the bottom layer is more saline than the top. Vertically homogeneous estuaries occur where tidal mixing is vigorous and freshwater input is low. In this case vertical salinity stratification breaks down.
The circulation patterns and physical characteristics of only a few Georgia estuaries have been studied. The Savannah River, a moderately stratified estuary, has been studied by the U.S. Corps of Engineers. It has a drainage area of approximately 9850 mile2 and an average discharge of 11,290 cubic ft per second (cfs) (U.S. Geological Survey 1960). It is tidal for approximately 50 miles upstream. A physical model of the river has been designed in order to study shoaling (U.S. Army Corps of Engineers 1949). Figure 30 shows the tidal currents at ebb and flood tides as predicted by the model.
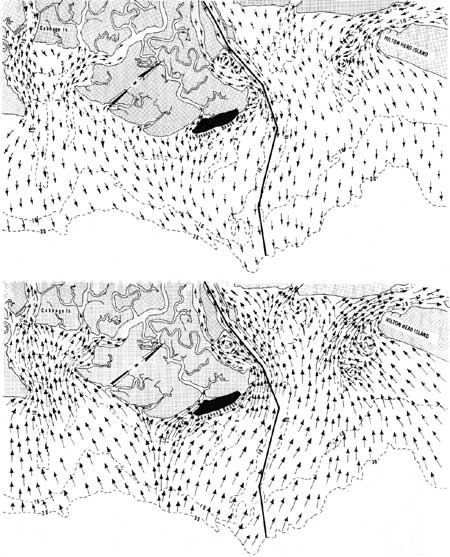
|
| Fig. 30. Direction of tidal currents at ebb and flood tides in the Savannah area as predicted by a physical model. (Redrawn from U.S. Corps of Engineers 1949). |
The Altamaha River has the greatest discharge along the Georgia coast. It has a drainage area of approximately 13,600 mile2 and an average discharge of 12,600 cfs (U.S. Geological Survey 1959). Several miles from the ocean it divides into a number of distributaries (Darien, Butler, Champney, and South Altamaha) which empty into Altamaha Sound and to a lesser extent into Doboy and St. Simons sounds. The hydrography of this river has been studied to only a limited extent. Neiheisel (1965), in a study of the causes of shoaling in Brunswick Harbor, presents information on the mixing and flow of water in Brunswick Harbor as influenced by the Altamaha River. Discharge into Brunswick Harbor, a well-mixed estuary, by the Turtle River is negligible. However, during maximum peaks of discharge, fresher water from the Altamaha enters the harbor through the Mackay and Frederica rivers. The water discharged through the Altamaha Sound into the ocean is moved along the shore and enters Brunswick Harbor through the sound entrance. Salinity measurements in the Mackay and Frederica rivers show that they are partially mixed estuaries with some density stratification during a period of low discharge. During higher discharge it is assumed that the interface would become sharper and move toward the harbor. Salinities were approximately 5 ppt in the Mackay River near Altamaha Sound and approximately 20 ppt near St. Simons Sound (Neiheisel 1965).
The hydrography of Doboy Sound, recently studied by Levy (1969), is also affected by the Altamaha River. During ebb tide, fresh water from the Altamaha flows seaward through the Darien-Rockdedundy complex into Doboy Sound through the Black, South, and North rivers. During mid-ebb this water flows out the mouth of Doboy Sound along the south shore, while water from the sounds leaves by the north shore. Because of the limited fresh water flow, Doboy Sound is a mixed estuary (at least during the summer months) with no recognizable salt wedge and little vertical stratification. A salt wedge is present just seaward of the inlet, where less saline, highly turbid waters of the longshore drift overlie more saline oceanic waters. This moves shoreward on flood tide.
Ragotzkie and Bryson (1955) studied the Duplin River, a tidal river located near Sapelo Island and opening into Doboy Sound. It has no significant fresh water source, and it is an elongated tidal bay about 5.9 miles long with a tidal excursion of about 3 miles. At mean low water the river has a relatively small horizontal area; however, at 6 feet above mean low water the water flows over the banks and onto the salt marsh. The area of marsh flooded increases rapidly as the level of water rises. From one-third to two-thirds of the volume of water entering the Duplin River on a rising tide leaves the river to flood the marsh. The volume of water involved is greatly increased with relatively small increases in tidal height. The waterways are subject to high turbulence as this large volume of water enters and leaves. Mixing is rapid and no stratification long can be maintained under these conditions.
Sediments
Coastal sediments have been the subject of considerable study since 1960. Much of this work was done by the University of Georgia Marine Institute. Papers dealing with sediments of the continental shelf (shelf sediments) and of the shallow waters near shore (paralic sediments) are reviewed by Henry and Hoyt (1968).
Previous sections of this report have described origins, composition, depositional patterns, and migrations of sediments of offshore bars, beaches, dunes, and marshes. The following summary is restricted to some ecologically important aspects of shelf and inshore sediments not previously discussed.
Nearshore, a narrow band of fine sands is considered to be of Recent origin (Pilkey and Frankenberg 1964). Minerals of mainland (terrigenous) origin (mainly quartz) are the dominant constituents (Henry and Hoyt 1968).
A relatively sharp boundary between Pleistocene and Recent sediments occurs quite consistently at about 6 fathoms, generally about 12 miles offshore (Pilkey and Frankenberg 1964). Cenozoic deposits on the continental shelf range from 2000 to 2500 ft deep (Henry and Hoyt 1968). Quaternary sediments consisting of coarse-grained particles are less than 100 ft thick and are mostly of Pleistocene origin (Henry and Hoyt 1968).
Carbonate contents of shelf sediments are discussed by Gorsline (1963), Pilkey (1964), and Pilkey et al. (1969). Carbonates average less than 25% over most of the shelf and consist mostly of mollusks. Carbonate content increases seaward to about 50% on the shelf edge and nearly 100% on the upper slope where foraminifera are the major components (Pilkey 1964). This increase toward the shelf edge probably results from the diminishing contribution of terrigenous materials.
The presence of oolites off the Georgia coast is indicative of pre-existing shoreline conditions and low sedimentation rates, and the abrasive history of the carbonate fraction indicates that former shorelines in this area, like those of the present, were subject to relatively low wave energies (Pilkey et al. 1969).
Phosphorite averages about 1.1% of shelf sediments (Henry and Hoyt 1968). Present-day rivers apparently lack phosphorite in their sediment loads, yet all shelf samples and most estuarine sands contain these grains in varying amounts (Pilkey and Terlecky 1966). Pevear and Pilkey (1966) and Pilkey and Terlecky (1966) concluded that much of the sands and nearshore fine sediments originate from landward transport from outer portions of the shelf and not from mainland rivers. Shelf phosphonte has evidently been derived from either Pleistocene rivers or from ancient phosphatic sediment outcrops on the shelf.
The heavy mineral fraction of shelf sediments averages less than 0.5% (Henry and Hoyt 1968). The heavy mineral suite includes staurolite, kyanite, garnet, zircon, epidote, pyroxenes, amphiboles, rutile, and tourmaline (Pilkey 1963). Heavy mineral distributions are frequently useful aids in the determination of sediment origins, but heavy minerals in shelf sediments off the Georgia coast show minimal variation because of their common origin in Piedmont rivers (Pilkey 1963).
Fauna of estuaries and inshore waters
Vertebrates
1. Fishes: The waters off the coast of Georgia support a variety of fishes related to the diversity of habitat. Some estuarine species enter fresh water to spawn and a few freshwater species enter the brackish estuaries. Some species are restricted to the estuaries and inshore waters and some are restricted to the waters of the continental shelf. But many species migrate between these habitats at various stages in their life cycles, and the estuaries are vitally important as nursery grounds and spawning grounds for many commercially important species harvested on the continental shelf. Stroud (1971) listed the species that are dependent upon the estuaries during some stage in their lives and reported that they comprised 63% of the Atlantic catch. He calculated that, for the Atlantic coast generally, each acre of estuarine habitat produces a yield of 535 lb on the continental shelf.
The continental shelf off Georgia generally is composed of shifting sediments and does not provide good fish habitat. However, a coral reef, or live bottom, recently has been discovered 16 miles due east of Cabretta Inlet on Sapelo Island. Such reefs provide a stable surface for the attachment of organisms important in the food chain. Artificial reefs also are being established by the Georgia Game and Fish Commission.
There has been relatively little work on the ecology of fishes of the Atlantic coast of the southeastern United States. Tagatz and Dudley (1961) studied the seasonality of fishes in four coastal habitats near Beaufort, N.C., and Tagatz (1968) and McLane (1955) surveyed the fishes of the St. Johns River, Fla.
Some pertinent work has been done off the Georgia coast. Anderson (1968) surveyed the fishes caught by shrimp trawling from South Carolina to north eastern Florida from 1931 to 1935. Miller and Jorgenson (1969) studied the seasonal abundance and length frequencies of fishes collected in two habitats and presented a list of fishes collected at a freshwater station in the Altamaha River. They made thorough surveys by seining at a beach habitat on St. Simons Island and two high marsh stations, one near Jekyll Island and one near Meridian, Ga. Dahlberg and Heard (1969) surveyed the common inshore elasmobranchs of the Georgia coast. Dahlberg and Odum (1970) sampled fish populations in St. Catherines and Sapelo sounds by trawling at 3-week intervals for 13 months. Struhsaker (1969) presented a list of fishes taken during 5 years of exploratory trawling on the continental shelf off Georgia and other southeastern states.
A list of fishes that may be encountered in Georgia's coastal waters is included (Appendix 1).
2. Reptiles: The loggerhead turtle, previously discussed, is the only marine reptile that is common on the Georgia coast. Green and Ridley's turtles have been reported from coastal Georgia (Appendix 2).
3. Marine mammals: Information on marine mammals of Georgia coastal waters is restricted mainly to stranding records and associated data. Only six species of whales, all toothed (Odontoceti), are recorded as having stranded on the island beaches, although sight observations and strandings from adjacent states indicate that about 23 species are probably components of Georgia's offshore fauna.
Atlantic bottle-nosed dolphins are probably the most common whales in the waters immediately adjacent to the coast. They are observed, frequently in pairs, swimming off the beaches and tidal rivers. Stranding dates are confined to the last half of the year, although the dolphin is commonly sighted throughout the year.
The pilot whale, also common, occasionally strands in large numbers on the island beaches. Between 15 and 25 stranded on St. Simons Island in 1962, and 53 stranded on Little St. Simons in 1968 (Caldwell et al. 1971). There is, as yet, no satisfactory explanation for these mass strandings.
Occasionally other whales such as the rare goosebeaked whale (with less than 20 known specimens from the western North Atlantic—Moore 1963) strand on the coast or are observed in the offshore waters.
The manatee occasionally is sighted or collected on the coast (Tomkins 1956, 1958), but its status as a permanent resident is doubtful.
Man inadvertently has introduced the California sea lion into the coastal waters of the southeastern United States, and sight observations (Caldwell et al. 1971) confirm the presence of the species off Sapelo Island.
Specific records of marine mammals in Georgia are given in Appendix 5.
Invertebrates
Most studies of estuarine invertebrates have dealt with those forms relating in a basic way to the estuarine food chain (plankton) and with species having commercial significance (shrimp, crabs, oysters).
Jacobs (1968) reported that Acartia tonsa, Pseudodioptomus caronaius, and Paracalanus parvus were the most common zooplankton (copepods) encountered in the Duplin River and Doboy Sound. Other copepods included Centropages hamatus, common in the spring, rare in summer, and Labidocera aestiva, common in the summer and absent during the spring. High concentrations of zooplankton often occur in the estuaries. Jacobs (1968) recorded 20,000 Acartia tonsa per cubic meter of water near Sapelo Island. Studies relating to the taxonomy, distribution, and abundance of estuarine invertebrates include the following: Frankenberg (1966), Nassarius trixittatus; Frankenberg and Menzies (1966), a new isopod; Marcus and Marcus (1967), opistobranchs; Maturo (1966) bryozoa; Menzies and Frankenberg (1966), isopods; Bullis and Thompson (1965), crustaceans and mollusks; and Frankenberg (1968), amphioxus.
The invertebrate fauna of Sapelo and St. Catherines sounds and adjacent tidal rivers recently has been sampled by Heard and Heard (1970). Fouling organisms, or those inhabiting solid substrates such as rocks and pilings, were studied by taking monthly samples from the bottom and sides of a floating dock at Colonel's Island (Liberty County). Forty-seven species or forms representing the following phyla were obtained: Porifera, Cnidaria, Ectoprocta, Mollusca, Annelida, Arthropoda, and Chordata. Table 14 is a listing of those species that were considered common or abundant by the investigators in at least one monthly sample. Benthic epifauna and infauna were sampled within the sounds using a 20-ft otter trawl and a bucket dredge. One hundred and eighty-five genera or forms were reported. Of these, 79 were considered common in at least one sample. These species are listed in Table 15. Classification or higher taxa have been changed in Tables 14 and 15 to agree with those of Meglitsch (1967). The restricted sample area and the selection by the collecting devices for certain invertebrates contributed some bias to the lists.
Most invertebrates of commercial importance (e.g., crabs, oysters, and shrimp) have been extensively studied. Following is a brief discussion of blue crabs, oysters, and brown and white shrimp.
Studies by Durant (1970) indicate that in Georgia, oysters (Crassostrea virginica) begin to spawn when the temperature is about 73° F. Spawning was observed to begin in May and to continue until October, with peak periods in July, August, and September (Durant 1970). Larval stages last for 2-3 weeks (Wallace 1966), after which the young attach to some substrate. Galtsoff (1964) states that only soft mud and shifting sand are totally unsuitable. However oysters may convert a mud bottom to a more suitable habitat if a few settle on a hard object and themselves become objects of attachment. Galtsoff describes the soft mud bottom of the South Atlantic as being only marginally suitable for oysters. He further states that oysters need a free exchange of water salinities of 5-30%, and temperatures from 34°F to 86°F. Conditions are ideal for feeding when the water, free of pollution and containing a low concentration of small diatoms and dinoflagellates, moves over the bottom in a nonturbulent flow.
The negative factors influencing oyster production are described by Wallace (1966) as "pollution, predators, and people." He reports that oyster production is inversely proportional to human population growth in New England and the mid-Atlantic states. Only in the southeastern and Gulf states does oyster production even approach that of 20 years ago. Wallace (1966) concludes that pollution is the primary cause of the decline of the oyster industry. Sewage is detrimental because it covers the bottom with sludge that smothers oysters and reduces oxygen (Galtsoff 1964). When Escherichia coli, bacteria associated with fecal matter and used as an index for pollution, reach certain numbers, the oyster grounds are closed for health reasons. Figures 35A-H (Appendix 6) show areas closed to oyster gathering by the Georgia Department of Public Health as of 1970. Industrial wastes also affect oysters. Galtsoff (1964) reports that red liquor and black liquor, both wastes from pulp mills, reduce the length of time the oyster shell remains open, thereby reducing the time available for feeding. Butler (1966) found that shell deposition is decreased in the presence of chlorinated hydrocarbon insecticides (e.g., DDT) at concentrations as low as 10 parts per billion (ppb). Oysters are especially susceptible to pollution because of their stationary mode of existence and their ability to concentrate pollutants in their tissues. Predators include flatworms, mollusks, echinoderms, crustaceans, fish, birds, and mammals (Galtsoff 1964).
TABLE 14. Epifauna collected from docks at Colonels Island (North Newport River, St. Catherines Sound).a
|
Phylum Cnidaria Unidentified anemone Unidentified hydroid Phylum Ectoprocta Anguinella palmata Alcyonidium sp. Amathia distans Conopium tenuissimum Phylum Annelida Class Polychaeta Nereis (Neanthes) succinea Polydora sp. Phylum Arthropoda Class Pycnogonida Tanystylum orbiculare Class Crustacea Subclass Copepoda Doropygus laticonis Order Thoracica Balanus eberneus Order Amphipoda Amphithoe valida Corophium simile Gammarus mucronatus Lumbos websteri Mellita appendiculata Paracaprella tenuis Parapleustes n. sp. Order Decapoda Paleomonetas vulgaris Paleomonetas pugio Eurypanopeus depressus Panopeus herbstii Phylum Chordata Subphylum Urochordata Molgula manbattensis |
aHeard and Heard 1970. |
TABLE 15. Invertebrate fauna of Sapelo and St. Catherines sounds.a
|
Phylum Porifera Halichondria sp. Microciona prolifera Phylum Cnidaria Leptogorgia virgulota Renilla reniformis Phylum Nemertina Bicolored form Phylum Phoronida Unidentified species Phylum Ectoprocta Aeverrillia setigera Alcyonidium verrilli Amathia distans Anguinella palmata Bowerbankia gracilis Bugula neritina Membranipora arborescens Membranipora tenuis Schizoporella unicornis Phylum Mollusca Class Gastropoda Anachis avara Busycon canaliculatum Busycon carica Cerithiopsis emersoni (C. subulata) Crepidula piano Doridella burchi Eupleura caudata Mitrella lunata Nassarius vibex Polinices duplicatus Pryogocythana (Mangelia) plicosa Terebra dislocata Turbonilla (Pyrgiscus) interrupta Class Pelecypoda Abra aequalis Amygdalum papyria Barnea truncata Ensis directus Lyonsia hyalina Mulinia lateralis Musculus lateralis Mya arenaria Solon viridus Spisula solidissima Tagelus divisus Tellina agilis Class Cephalopoda Lolliguncula brevis Phylum Annelida Class Polychaeta Arabella iricolor Clymenella torquata Diopatra cuprea Glycera americana Harmothoe-Lepidonotus spp. Heteromastis sp. Hydroides dianthus Lumbrineris tenuis Marphysa sanguinea Nereis (Neanthes) succinea Notomastis sp. (spp?) Parapironospio pinnoto Pectinaria sp. (gouldii?) Sabella microphthalma Sabellaria vulgaris Sabellaria sp. (floridensis) Sabellides sp. (oculatus?) Scoloplos-Orbinia spp. Spiochaetopterus oculatus Sthenelais boa Phylum Arthropoda Class Pycnogonida Anoplodactylus sp. Class Decapoda Acetes americanus carolinae Alpheus normanni Eurceramus praelongus Latreutes parvulus Lysmata wurdemanni Pagurus longicarpus Pagurus pollicaris Palaemonetas pugio Palaemonetas vulgaris Penaeus aztecus Penaeus setiferus Periclimenes longicaudatus Trachypenaeus constrictus Upogobia affinis Phylum Echinodermata Class Asteroidea Asterias forbesi Luidia clathrata Class Ophiuroidea Hemipholis elongata Ophiothrix angulata Phylum Chordata Subphylum Urochordata Molgula manhattensis aHeard and Heard 1970. |
The predominant species of marine shrimp occurring in Georgia waters are the white shrimp (Penaeus setiferus) and the brown shrimp (P. aztecus), both of which are important commercially. The life cycles of white and brown shrimp are basically similar. The bottom-dwelling (benthic) adults release their eggs freely into the waters offshore. Within a short time, the eggs hatch into planktonic larvae. After passing through several intermediate stages, the young shrimp (postlarvae) move into the estuary and adopt a benthic existence (Anderson 1955). After very rapid growth, they assume the adult form. Marking studies indicate that after migrating offshore the shrimp do not move into deep water but make seasonal migrations parallel to the shoreline (Anderson 1955). White shrimp penetrate the estuary to a greater degree, arrive later, and stay for a longer period of time than the brown.
Salinity optima for young penaeid shrimp are in the range of 5-20%, although shrimp can tolerate salinities from 1 to 600% (Kutkuhn 1966). A complex interaction of factors including circulation, temperature, salinity, and fertility of waters and type of vegetation and substratum determines distribution, survival, and growth of young shrimp (Kutkuhn 1966). Optimum conditions are approached in the nursery grounds of the marsh-estuary complex.
Nichols and Keney (1963) report that the identity and distribution of crabs of the genus Callinectes on the southeastern coast of the United States is uncertain. Rathbun (1930) reported two species, C. sapidus and C. ornatus, occurring between New Jersey and Indian River Inlet, Fla. Lunz (1958) found that only 30% of the crabs caught by trawlers in South Carolina were C. sapidus. The two species are not recognized as such by fishermen and are combined as blue crabs in catch data reported for coastal Georgia.
Van Engel (1958) reports that in the Chesapeake Bay area Callinectes sapidus begins mating early in May and continues into October. Females probably mate only once, at the time of the last molt. Sperm live in the female receptacles for at least a year and may be used as often as the female spawns (two or more times). The females migrate to saltier waters after mating, some passing out of the bay and into the ocean. Spawning is delayed at least 2 months after mating. When laid, the eggs are attached to the abdomen of the female where they remain about 2 weeks until hatching. Van Engel (1958) reports that there are two larval stages, four or five zonal stages, and the megalops. These stages are passed through in about 1 month, after which the first crab stage is reached. Costlow and Bookout (1959) observed seven zonal stages in laboratory-reared animals. Nichols and Keney (1963), based on the occurrence of early stage larvae, believe that spawning occurs throughout the year. Peak numbers of first-stage larvae were found in Georgia waters during July, August, and September, and large numbers of first and second stage zoeae were found near the beaches with progression to advanced stage zoeae 20-40 miles offshore. Van Engel (1958) reported that early in August many crabs reach the "first crab" stage and begin migrating into waters of lower salinity. Male crabs remain in less saline waters year round.
Thus blue crabs are a part of both the benthic and planktonic communities, and they use both inshore and offshore waters.
Fishery resources of estuaries and inshore waters
The estuarine and inshore waters of Georgia support a moderate-sized commercial fishery. The commercial harvest of fish and shellfish for the period 1960-65 averaged 21.4 million pounds valued at approximately $3.4 million (Carley 1968). The catch accounted for about 15% by weight and 17.6% by value of the harvest of the South Atlantic states. The commercial harvest of fish and shellfish has fluctuated considerably over a period of years, but total monetary value of the catch has been relatively stable because of increasing prices.
The shrimp fishery is the most important commercial fishery in Georgia, accounting for 83% of the harvest value (Carley 1968). Blue crabs account for one-half to three-fifths of the pounds of the annual harvest but account for less than 10% of the harvest value (Carley 1968). Oysters average about 1% of the total catch. Shad are the most important fish in the catch and, with other fish species, make up approximately 5% of the total catch (Carley 1968).
Shrimp are harvested on the Georgia coast by trawling during the 6-month season from 1 June - 31 December. The area open to harvest extends from the mouth of the sounds to 3 miles offshore. Shrimping outside the 3-mile limit is legal throughout the year. White shrimp dominate the catch until late in July when they are replaced by the brown shrimp. The browns then dominate the catch until they are again replaced by white shrimp late in August. The catch of white shrimp increases until it peaks in October. The total catch normally consists of two-thirds white shrimp and one-third brown shrimp (Carley and Frisbie 1968a).
Trawling is legal in Georgia during the daytime only. This regulation is a significant factor influencing the composition of the total catch. The white shrimp feeds during the day and burrows into the mud at night. Conversely, the brown shrimp is nocturnal, burrowing into the mud during the day.
The pink shrimp (Penaeus duorarum), also commercially important in some areas, occurs infrequently in the catch in Georgia. This shrimp has habitat requirements (relatively clear, shallow waters with a firm bottom) that are met in relatively few areas in Georgia waters.
Linton (1970) conducted a survey of the distribution and abundance of oyster beds in 1966 and 1967 and found about 10,180 acres of oyster beds, almost all of them intertidal and landward of the barrier islands. Oyster beds are better developed in the northern portion of the state where they occur in a band 7-8 miles wide (Figs. 35A-H, Appendix 6). All oysters in Georgia are taken from privately held areas through leasing arrangements (Carley and Frisbie 1968b). The most important factors limiting production are pollution, unsuitable substrate, and predation (Carley and Frisbie 1978b). Between one-third and one-half of the estuarine area suitable for oyster growth is polluted (Figs. 35A-H), and the oyster beds are closed to harvest for public health reasons (Carley and Frisbie 1968b). Little effort has been made to manage oyster beds on a long-term basis. Assuming effective pollution control were possible, oyster production could be significantly increased. Oyster shells and other material suitable for spat-setting could be deposited in many areas, and raft or line culture would be feasible in many areas (Shaw 1968).
A relatively small hard clam fishery existed in coastal Georgia until about 1932. A recent study of Godwin (1967) revealed in the Altamaha Sound a population of brackish water clams (Rangia cuneata) that could support a commercial fishery, but the area is closed for shellfish harvest because of pollution. The hard clam or quahog (Mercenaria mercenaria) does not occur in harvestable numbers in Georgia (Godwin 1967).
Blue crabs are taken in greater numbers than any other commercial species but, due to a low unit price, they represent a small portion of the monetary value of the catch (Carley 1968). Crabs are harvested with pots and traps and are taken in otter trawls incidental to shrimp trawling.
Finfish have averaged about 5% of the total quantity and value of all fish and shellfish in Georgia. Harvest by species is shown in Table 16. The greatest proportion of the catch of finfish in 1967, 36% of the poundage and 45% of the total economic value, was American shad (Lyles 1969). Shad are anadromous fish, spawning in fresh water and moving into the ocean to mature. The adults move up the rivers from January until April. Many shrimp and crab fishermen supplement their income by fishing for shad during this time. Young shad begin to move down the rivers in July and are found in the Atlantic by October (Godwin and Adams 1969).
Whiting made up 20% of the finfish poundage and 15% of the dollar value in 1967 (Lyles 1969). They usually are caught while trawling for shrimp. Carley (1968) reports that there has been a downward trend in the abundance and catch of whiting. There are three species of whiting inhabiting the coast of Georgia (Menticirrhus americanus, M. saxatilis, and M. littoralis), but they are not distinguished in commercial records. Menticirrhus littoralis prefers shallows and surf, so it is not often caught in trawls. Both M. americanus and M. saxatilis spawn in spring and early summer. The young of the former occur in both offshore and inside waters, whereas those of the latter species live in the surf. The adults of both species are found in both inside and outside waters (Hildebrand and Cable 1934).
Red snapper accounted for 6% of the poundage and 17% of the cash value of the 1967 catch, and grouper accounted for 12% and 10%, respectively. These are by far the largest catches reported for these two species since 1957 (Power 1959; Power and Lyles 1964; Lyles 1965, 1966, 1968, 1969). Both species are caught with hand lines off the live bottom and shelf edge habitats. Red snapper are also caught in trawls (Lyles 1965, 1969; Struhsaker 1969).
The total value of the commercial fishery is probably underestimated. Carley (1968) reported that in many cases fish caught while shrimping were given to the crew of the vessel and were not reported. Menhaden form the base of the largest fishery in the United States by volume. The larvae and juveniles spend their first summer in the estuaries where they grow rapidly and serve as an important food item for many carnivorous species. They move into deeper waters in the fall where they are caught in large purse seines (June 1961). Approximately 2000 purse-seine sets were made in Georgia in 1959, each averaging 20-25 tons of fish (June 1961): a total of 40,000 tons. Because there are no menhaden processing plants in Georgia, these fish were landed outside the state, probably at Fernandina Beach, Fla., and are not listed in the Georgia landings.
Sport fishing is also an important element in the coastal economy. Expenditures by an estimated 281,400 salt water sport fishermen on the Georgia coast totaled $22,523,280 in 1968 (Cheatum et al. 1968). Table 17 lists the sport fish most commonly caught on the Georgia coast. Harvest figures are not available for salt water fish taken by sport fishermen in Georgia.
TABLE 16. Fish with total landings in Georgia of more than 000 lb per year.a
| Species | Pounds | |
| 1966 | 1967 | |
| Croaker | 5,100 | 6,000 |
| Flounder | 33,500 | 22,000 |
| Whiting | 145,400 | 186,700 |
| Sea bass | 2,700 | 1,700 |
| Black drum | 2,400 | |
| Red drum | 5,600 | |
| Grouper | 92,300 | |
| Red snapper | 54,900 | |
| Weakfish | 1,300 | |
| Spotted sea trout | 1,200 | 6,900 |
| Spanish mackerel | 1,300 | 2,000 |
| Spot | 5,200 | 10,500 |
| Bait and animal food | 127,600 | 203,400 |
| Hickory shad | 2,000 | 1,300 |
| American shad | 385,900 | 334,100 |
| Mullet | 2,000 | |
aLyles 1968, 1969. | ||
TABLE 17. Common salt water sport fishes of Georgia
| Inshore species | Offshore species |
| Spotted sea trout | king mackerel |
| Red drum (channel bass) | dolphin |
| Spanish mackerel | little tuna |
| Tarpon great | barracuda |
| Bluefish | Atlantic bonita |
| Cobia | pompano |
| Sheepshead | crevalle jack |
| Striped bass | greater amberjack |
| American shad | red grouper |
| Southern kingfish | red snapper |
| Triple tail black | sea bass |
| Southern flounder | sailfish |
| Weakfish | |
| Atlantic croaker | |
| Spot | |
Productivity and energy flow
Nutrients
The waters comprising the coastal region are actually a solution of many inorganic and organic compounds and other materials in suspension. The movements and concentrations of these substances are important aspects of the ecology of the area. Elements such as carbon, nitrogen, and phosphorous are necessary for primary production and often prove to be limiting factors. Of these, phosphorus and zinc have been studied in the Georgia coastal waters.
Phosphorus is often limiting in oceanic waters (Raymont 1963). Pomeroy et al. (1965) found phosphorus concentrations of 1 micromole per liter (µM/L) in Doboy Sound. At this high concentration phosphorus was not considered to be a factor limiting primary production. Estuarine water of the Altamaha River contains phosphorus in concentrations ranging from 0.05 to 4.0 µM/L, whereas the river water has a phosphate concentration of 0.1 µM/L. Pomeroy et al. (1969) concluded that the estuary acts as a "sink" since it accumulates sediments that contain organic matter as well as many minerals. Experiments have been conducted in which 32P was released in the headwaters of the Duplin River (Pomeroy et al. 1969). From this work it was determined that the sediments play an important role in the phosphorus cycle. Phosphorus from the surface sediments, which are in equilibrium with the water, moves into a number of deposit feeders and from them, via excretion, back into the water. Subsurface sediments, not in equilibrium with the water, are the source of phosphorus for smooth cordgrass. Bacteria within the sediments are probably accumulating phosphorus, which is released by the bacterial feeders (Johannes 1965). Thus, there is a separation between pathways within the salt marsh. Phosphorus in subsurface sediments is incorporated into the cordgrass which serves as the principal source for insects and spiders. Most of the cordgrass, however, dies and enters the detritus food chain. Thomas (1966) showed that the estuarine waters, rich in phosphorus, are flushed out into the offshore waters and serve as a nutrient source for phytoplankton.
Zinc is a trace element required by some organisms and often is concentrated by them. Tracer studies with 65Zn indicate that the cycle of zinc, like that of phosphorus, is dominated by the sediments which act as a mixed-bed ion exchanger (Pomeroy et al. 1969). Organisms, except fiddler crabs and oysters, take up zinc and phosphorus at much the same rates. Oysters demonstrate a preferential retention of zinc, and fiddler crabs show high initial values of zinc activity, falling off rapidly and becoming erratic, suggesting movements in and out of the area. Bioelimination of zinc seems to be slow. Concentrations of zinc are, like phosphorus, high in the Duplin River as compared to sea water, and the same processes that concentrate phosphorus may concentrate zinc (Pomeroy et al. 1969).
Vitamin B12 is a nutrient essential for growth of many organisms such as bacteria, algae, and protozoa. It is synthesized mainly by bacteria and fungi (Burkholder and Burkholder 1956) and is found in large quantities in marine invertebrates and fish. Starr (1956) and Burkholder and Burkholder (1956) studied the distribution of this vitamin in suspended solids and marsh muds from the Georgia coast. Burkholder and Burkholder (1956) found that the dark-water rivers contained high concentrations of B12 per unit of suspended particles. Both studies showed a progressive increase in B12 from offshore stations to the head of a tidal creek. However, Starr (1956) reports that ocean waters contain detritus richer in B12 than the more turbid sound waters. The highest concentration of B12 is contained in the detritus at the headwaters of the tidal rivers. Starr (1956) also established that detritus entering the marsh at high tide was not as rich in B12 as that leaving on the ebb tide. Sediment samples taken from areas devoid of marsh were much lower in B12 than those taken from marsh areas; however, both were lower in B12 than water leaving the marsh. Starr (1956) concluded that the muds did not add appreciable quantities of vitamin-rich detritus to the waters. Burkholder and Burkholder, however, report that muds treated with sulfite yield values 3.8 times higher than untreated samples, and Starr did not mention this technique which could have changed his conclusion. More work is needed to clarify this problem as well as the role of B12 in productivity of the sea.
Most of the turbidity of coastal waters is caused by suspended sediments, but a significant portion (18%) is caused by organic material (Odum and de la Cruz 1967). A standing crop of 2-20 micrograms ash-free, dry organic matter per liter is reported for the Georgia estuaries, a value much greater than reported for open sea water or other sounds and bays (Odum and de la Cruz 1967). These measurements were taken at the mouth of a small tidal creek draining a salt marsh. Samples taken at mid-ebb tide always produced more organic detritus than did samples taken at mid-flood tide. This suggested that organic material is being exported from the marsh. Teal (1962) also calculated a net flow of organic material from the marsh to the estuary and beyond.
The waters of the coastal region appear to be rich in organic materials, the two inorganic nutrients phosphorus and zinc, and vitamin B12. All of these are found in higher concentrations in the salt marsh-estuary system than in the open ocean.
Food webs
Food webs of the estuary are not nearly as well understood as those of the salt marsh. Many of the conclusions concerning the food webs of the Georgia estuaries must be inferred from work done in other regions. The estuarine environment is more complex than the marsh system, and many estuarine species migrate seasonally. Estuarine inhabitants can be categorized as plankton, benthic infauna, epifauna or fouling organisms, and nekton. Some are deposit feeders, some suspension feeders, and some are carnivores.
Specific food habits studies have been conducted on commercially important invertebrates such as crabs, shrimp, and oysters. Shrimp are deposit feeders; Darnell (1958) reports that detritus and fine organic material composed 58% of the food of shrimp examined, and mollusks 12%. He suggested that both young and mature white shrimp are omnivorous, feeding largely on detritus. Oysters are suspension feeders. Galtsoff (1964) studied ciliary currents produced by oysters and concluded that the food-sorting mechanism is designed for continuous feeding in low concentrations and that food selection, except on the basis of size, is open to question. The blue crab is both a scavenger and a predator; foods include mollusks, detritus, crabs (Callinectes, Rithropanopeus, and unidentified species), vegetation, and fish remains (Darnell 1958).
Data from published studies of the stomach contents of fish (Table 18) are summarized in Table 19 to show generalized feeding habits of estuarine fishes. Food items are divided into the following general types: detritus, algae, zooplankton, microbenthic invertebrates, macrobenthic invertebrates, shrimp, and fish. The majority of fish consume more than one type of food and are not confined to one trophic level. Ninety-two percent of the fish fed to some extent on benthic invertebrates. These percentages are based only on those common species which have been studied, and the data were collected with varying degrees of precision from widely scattered areas. Some of the studies took the age of the fish into account; others did not. However, in every case where stomach analyses were done for individual size classes, a change of food habits was correlated with a change in size. For example, croaker less than 25 mm in length eat principally zooplankton; those 25-50 mm feed mostly on microbenthic in vertebrates. Detritus, shrimp, and macrobenthic invertebrates are the main food items of fish 50-200 mm. Large croaker (greater than 200 mm) eat macrobenthic invertebrates, shrimp, and fish (Darnell 1958).
TABLE 18. Some food habits studies of common estuarine fishes.
| Species | Reference |
| Sphyrna tiburo (bonnethead) | Gunter 1945 |
| Lepisosteus osseus (longnose gar) | Goodyear 1967 |
| Megalops atlantica (tarpon) | Rickards 1968 |
| Bevoortia sp. (menhaden) | June 1961 |
| Anchoa mitchilli (anchovy) | Darnell 1958 |
| Galeichthys felis (sea catfish) | Darnell 1958 |
| Opsanus tau (oyster toadfish) | Schwartz and Ducher 1963 |
| Pomatomus saltatrix (bluefish) | Grant 1962 |
| Orthopristis chrysopterus (pigfish) | Hildebrand and Cable 1934 |
| Archosargus probatocephalus (sheepshead) | Gunter 1945 |
| Lagodon rhomboides (pinfish) | Darnell 1958 |
| Bairdiella chrysura (silver perch) | Darnell 1958 |
| Cynoscion nebulosus (spotted sea trout) | Darnell 1958 |
| Cynoscion regalis (weakfish) | Hildebrand and Cable 1934 |
| Leiostomus xanthurus (spot) | Darnell 1958 |
| Menticirrhus americanus (southern kingfish) | Gunter 1945 |
| Menticirrhus littoralis (Gulf kingfish) | Gunter 1945 |
| Menticirrhus saxatilis (northern kingfish) | Welsh and Breder 1923 |
| Micropogon undulatus (Atlantic croaker) | Darnell 1958 |
| Pogonias cromis (black drum) | Welsh and Breder 1923 |
| Sciaenops ocellata (red drum) | Darnell 1958 |
| Stellifer lanceolatus (star drum) | Welsh and Breder 1923 |
| Mugil cephalus (striped mullet) | Darnell 1958 |
| Paralichthys lethostigma (southern flounder) | Darnell 1958 |
| Chilomycterus schoepfi (striped burrfish) | Gunter 1945 |
TABLE 19. Feeding habits of estuarine fishes.a
| Type feeder | Per cent of species studied |
| Planktonic feeders only | 4 |
| Benthic feeders only | 32 |
| Fish feeders exclusively | 8 |
| Detritus feeders only | 0 |
| Plankton feeders | 44 |
| Benthic feeders | 92 |
| Fish feeders | 52 |
| Detritus feeders | 16 |
| Strict herbivores | 0 |
| Strict carnivores | 34 |
| Shrimp feeders | 52 |
| Limited to one type food | 28 |
| Eat more than one type food | 72 |
| Eat more than two types of food | 52 |
| Eat more than three types of food | 32 |
| Eat more than four types of food | 12 |
aSummarized from data reported from studies listed in Table 18. | |
| <<< Previous | <<< Contents >>> | Next >>> |
chap5.htm
Last Updated: 1-Apr-2005