
|
Geological Survey Professional Paper 294—D
Stromatolites of the Belt Series in Glacier National Park and Vicinity, Montana |
ECOLOGY
Stromatolites occur in rocks ranging in age from Precambrian to Recent. Recent occurrences provide a basis for interpreting the conditions favorable for their development. As stromatolites are not the actual remains of algae but structures that were developed by the reactions of the organisms to their physical environment, they are, in effect, a reflection of the environment in which they developed. The physical laws that govern the reactions of matter to changes in temperature, salinity, moisture content, and other conditions presumably operated during the Proterozoic in the same manner as they do today. Thus, although many of the modern stromatolite-forming algae are able to exist under a wide range of environmental conditions, others are restricted to certain combinations of environmental factors. In the following discussion an attempt will be made to lay the groundwork for interpreting the possible conditions of origin of the stromatolites studied—it is not intended, of course, to set forth strictly limiting environmental factors for each species.
MODERN ENVIRONMENTS
Two areas in which modern stromatolites have been reported to be developing in marine and near-marine environments are southern Florida (Ginsburg,2 1954, 1955) and Andros Island, Bahama Islands (Black, 1933). During the fall of 1954 and the spring of 1955, I visited these areas in order to obtain firsthand information on the occurrences.
2Ginsburg, R. N., and others, 1954, Laminated algal sediments of south Florida and their recognition in the fossil record: Univ. Miami Marine Lab., Coral Gables, Fla., unpub. manuscript.
Maurice Black (1933), in his paper on the algal sediments of Andros Island, describes in detail the distribution of marine algae on and near that island. He also comments on the similarity of the modern stromatolites to structures existing in Precambrian and Paleozoic sedimentary rocks. The following table lists the four algal zones on Andros Island as reported by Black.
Algal zones on Andros Island [After Black, 1933]
| Maximum salinity (parts per thousand) |
Nature of habitat | Flora |
| 30-40 | Shoal water below tides | Chlorophyceae: Udotea, Penicillus, Halimeda. |
| 15-36 | Between tides near high water-mark. | Cyanophyceae: Phormidium, Symploca, and unicellular forms. |
| Trace-2 | Above high watermark, inland marl date. | Cyanophyceae: Scytonema, Pleotonema, Schizothrix, and unicellular forms. |
| 0 | Above high watermark, limestone outcrops. | Scytonema alone. |
In the shoal-water habitat, Black observed no complex algal heads, and the Cyanophyta which are responsible for these structures are not present in large numbers. The predominant algae in this habitat are the Chlorophyta, which bind the sediment without producing any laminated structure. Black remarks (1933, p. 169) that it was only above low water mark, where the Cyanophyta are the prevalent algae, that the alga controlled lamination and algal heads with characteristic internal structures were found.
W. R. Taylor (1928) does not report any stromatolites in his paper on the marine algae of Dry Tortugas. However, he refers to a total of 42 species of Cyanophyta, of which 31 are littoral or sublittoral. Concerning the littoral zone, he states (1928, p. 34), "the Myxophyceae (Cyanophyta) constitute an important part (31 forms) and this zone is the only one in which they are significant."
M. A. Howe described the algal flora of Bermuda in 1918. His comments on the occurrence of Cyanophyta (1918, p. 491) bear out the observations of Black and Taylor.
The Cyanophyta may form gelatinous or slimy dark scums on rather stagnant water or somewhat similar films or crusts on rocks or on the larger aquatic plants, either between the tide lines or permanently submerged. They are not confined to the sea, but are perhaps even more common in brackish or fresh water. They also occur on moist ground and even on the trunks of trees and rocks where rains and atmospheric moisture are their only sources of water supply.
My own observations of the distribution of Cyanophyta along the seashore are in agreement with those of Black, Taylor, and Howe. Marine Cyanophyta are most abundant above low watermark. In this zone, only those algae capable of surviving long periods of drought and considerable changes in salinity are able to exist. It is on the moist ground and rocks in and above this zone that stromatolites are most likely to develop.
Black described 4 types of algal deposits on Andros Island. All these deposits were found developing above low watermark. In southern Florida, Ginsburg3 reports 2 types of algal deposits that coincide with 2 of Black's types. The simplest, type A of Black and type I of Ginsburg, occur in areas frequently inundated by seawater (intertidal zone near high watermark). Filamentous blue-green algae permeate the sediment as it is deposited and bind it together. At times of non-deposition, a thin algal mat is formed. The alternation of these organic layers merely accentuates the mechanically formed laminae. The alga does little else to modify the sediment. (See fig. 51.)
3Op. cit.

|
| FIGURE 51.—Algal deposits of type A (X 2) (after Black, 1933). |
The sediment is composed of two types of laminae. The thicker laminae are between 0.5 millimeter and 2.0 millimeters thick and are made up of detrital grains that are up to 0.1 millimeter in diameter. Interbedded with these distinctly sedimentary laminae are thinner layers, about 0.1 millimeter thick, of very fine-grained sediment. These layers are apparently made up of the grains that were entrapped in the mucilaginous sheaths of the algae. Some of the finely laminated sedimentary rocks in the Belt series could have developed in a similar manner.
Black's type B is similar to one of the forms described by Ginsburg under his type II. These deposits are formed above high watermark beyond the immediate influence of shore flooding. (See fig. 52.) They are irregularly disposed hollow domes or bubbles, 1 or 2 inches in height and 4 or 5 inches in diameter. They exhibit crude concentric lamination that is due to the differences in grain size in the successive layers and to the alternation of light and dark layers in which the grains are imbedded in the remains of algae. Ginsburg4 mentioned that reflooding of the area destroys the domes and that they are only seasonal features. However, distortions in the subsurface laminae are probably due to old domes. Bubblelike distortions have been observed in finely laminated sedimentary rocks of the Belt series and later deposits.
4Op. cit., p. 21.

|
| FIGURE 52.—Algal deposits of type B (X 1) (after Black, 1933). |
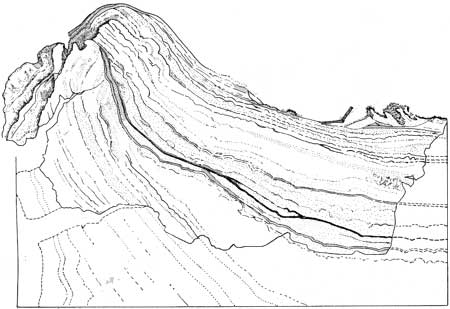
According to Black, the most highly developed types of algal head and the most extensive algal deposits are found bordering the fresh-water lakes in the interior of Andros Island. Along the margins of these lakes are large areas of limy mud marshes that are covered by algal heads that exhibit nearly concentric laminae. Deposits of type C appear as regularly spaced raised discs, from 4 to 5 inches in diameter. Black (1933, p. 173) remarks:
This growth form is the result of two opposing tendencies. The growth process, by itself, gives rise to flat or smoothly convex forms similar to those of type B; this simple shape, however, is rarely retained for long and is constantly liable to modification during periods of partial drying. Owing to the large proportion of organic matter, and fine texture of the sediment in the algal heads, drying causes the upper layers of the sediment to contract and a concave shape results. [See fig. 53.] Renewed growth of the algae produces further layers, at first concentric with the older part of the algal head, but gradually re-establishing the Convex form again. [See fig. 54.]
|
| FIGURE 53.—Algal deposits of type C (X 2) (after Black, 1933). |
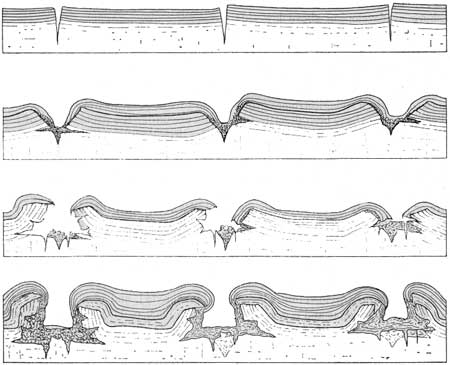
|
| FIGURE 54.—Sequence of growth of algal deposits of type C (X 1/4) (after Black, 1933). |
Three genera and three or four species contribute to the formation of these structures. The sediment consists of angular detrital grains of calcite averaging about 4 microns in diameter.
Deposits of type D are similar to the earlier formed parts of algal heads of type C except in their having a greater proportion of organic material to sediment and in the less orderly form of their laminae. During dessication they roll up and become completely detached from the substratum. (See fig. 55.) The algae of type D deposits are the same as those in type C but differ in proportion. The sediment is extremely fine grained; most particles are 2 microns or less in diameter. According to Black, this material is a limy mud that has been washed into the interior of the island by hurricane waves. Except for these occasional inundations of the island's interior by sea water, the algal heads in that area are soaked by rainwater and fresh ground water containing large amounts of dissolved calcium carbonate.

|
| FIGURE 55.—Algal deposits of type D (X 2) (after Black, 1933). |
Structures similar to Black's types C and D also occur in southern Florida. Ginsburg has treated these as variants of his type II. In addition to the stromatolites described by Ginsburg as developing above low tide level, he cites (1955, p. 724) the occurrence of small discrete spheroidal forms in Florida Bay at depths of 1 to 6 feet. These spheroidal forms are unattached and free to roll about on the bottom. These are onkolites similar to Ottonosia Twenhofel from the lower Permian.
Several occurrences of modern spongiostromes growing in continental environments have been described in the published record (Bradley, 1929; Clarke, 1900; Howe, 1932; Jones, 1925; Pia, 1933; Roddy, 1915; Walcott, 1914). Bradley (1929, p. 204-206) describes deposits along the shores of Green Lake, N. Y. He states, "All these deposits are exceedingly porous or spongy and consist of more or less closely intergrown arborescent masses that are richly nodose. They resemble native copper but are more profusely branched." Here, the form of the deposit is controlled by the growth habit of the algae, and it can be readily comprehended that a fine precipitate of lime is deposited about the filaments of the algae. This deposit of finely divided lime adheres to the mucilaginous sheath of the algae and is gradually built up into a spongy mass. (See pl. 24, figs. 3 and 4.) Bradley's descriptions do not fit any of the stromatolites of the Belt series.
In 1900 Clarke (1900, p. 195) published a brief paper on the water biscuits of Squaw Island, Canandaigua Lake, N. Y. He states:
The north shores of Squaw Island and the lake bottom about it and over its northward sand bar are covered with flat, whitish calcareous cakes of circular or oval shape, in size ranging from a dime to a half dollar. To pick up one of these, well dried on the surface of the island, and break it in half, seems enough to convince the reflective mind at once of their nature and mode of formation. It often contains as a central nucleus a beach pebble of shale or limestone, a twig, or a bit of charcoal from some youngster's campfire. About this a white or greenish travertine has been deposited in concentric layers, which show themselves with distinctness. The calc-carbonate being dissolved in dilute acid and entirely removed, there remains a soft spongy organic residuum of precisely the volume of the original biscuit.
These concretionary masses grew unattached on the floor of the lake and belong to the group called Oncolithi by Pia (1927, p. 37). (See p. 129.) These masses, also, are unlike the stromatolites of the Belt series.
In 1915 Roddy (1915, p. 247) described calcareous concretions similar to those of Squaw Island from Little Conestoga Creek, Lancaster County, Pa.
Later, Howe (1932, p. 61) described similar concentrically laminated calcareous pebbles from Furnace Creek, a tributary of the Potomac River about 1-1/2 miles above Harpers Ferry, W. Va. Howe (1932, p. 62) states, "The first microscopic examinations of ground sections and of decalcified preparations showed a mixture of minute plants—diatoms, unicellular Chlorophyceae, unicellular and filamentous Myxophyceae [Cyanophyta], and possibly bacteria."
Pia (1933, p. 172) reports that algal crusts of the Bodensee [Lake Constance], Bavaria, yielded 12 species in 7 genera of green and blue-green algae and several species of diatoms. He states that there are many places on the earth where limy masses are originating through the activity of algae and these are identical with fossil forms called Cryptozoon, Pycnostroma, and others.
The monograph by I. C. Russell (1885) on the geologic history of Lake Lahonton includes several illustrations of calcareous tufa that apparently developed along the ancient shorelines. Several specimens, from recent collections in that area, have been supplied to me by Roger Morrison of the U. S. Geological Survey. Russell describes three types of tufa (1885, p. 190): "(1) Lithoid tufa; (2) Thinolitic tufa; (3) Dendritic tufa." Of these, only the thinolitic tufa appears to be of inorganic origin.
Some of the lithoid tufa is similar in structure to some of the Precambrian stromatolites. (See pl. 22, figs. 1 and 2.) The dendritic tufa resembles modern structures in Green Lake, N. Y. (See pl. 23, figs. 2 and 4.) One of the specimens from Morrison's collection is a fragment of a Recent algal mat from the shores of the Carson Sink, Nev. The mat consists of intertwined algal filaments that bind together a layer of detrital sediment. (See pl. 24, figs. 1 and 2.) Morrison states (oral communication, 1954) that the mat developed along the shores of the lake around 1860, when the level of the lake was considerably higher than at present. The level of the modern Carson Sink is quite low, and at times it dries up completely. The mat has been completely dessicated, and attempts to grow algae by placing fragments in water have been unsuccessful. According to Morrison (oral communication, 1954), such algal mats are at present forming deposits of lithoid tufa at Soda Lakes, about 9 miles northwest of Fallon, Nev. These deposits are very close to the present surfaces of the lakes and apparently are the only type of algal deposit forming at the present time. Perhaps the highly saline and alkaline composition of the lake water does not permit the growth of species that develop the dendritic tufa.
J. C. Jones (1925, p. 7) studied the origin of tufas in the Salton Sea and arrived at the conclusion that the tufas are the result of algal activity. Jones carried his study of modern tufas to Pyramid Lake, another remnant of Lake Lahonton, and found that tufas are forming there at the present time. The tufa consists of a thin layer covering the exposed surfaces of about half of the rocks and gravels along the shores. This thin layer extends from an elevation of 12 feet above lake level to an unknown depth beneath the surface of the lake. Jones (1925, p. 9) describes the distribution of algae in the lake as follows:
At the water's edge a long filamentous green alga, a species of Cladophora, grows as a fringe 6 to 9 inches long in a narrow band about equally divided above and below the water. When the level of the lake falls, this dries, bleaches, and forms a grayish-white, paper-like band clinging to the rocks. * * * Below this band a grayish-white alga, another species of Cladophora, with much shorter filaments, grew in tufts and bunches that had their bases so firmly fixed in the layer of recent tufa that pulling the algae from the rocks brought the tufa layer with them. At the base of the tufts was a narrow zone of blue-green algae that included species of Callothrix, Nostoc, and Formidium. * * * at depths of 4 feet and over, the blue-greens grew in the absence of the other algae, and in every instance in constant association with the recent tufa.
Jones further states (1925, p. 9) that the recent tufa forms a film up to 2 or 3 millimeters thick on the surfaces of rocks—"where the deposit is thin it can be seen to begin as a multitude of separate rounded points that rapidly increase in diameter, meet, and coalesce. As the deposit thickens it becomes solid, compact, and stony, and is a typical lithoid tufa."
Two distinct types of fossil lithoid tufa occur in the Lake Lahonton region. The first is a dense, compact, laminated structure, irregularly shaped, that appears to have been formed under water. This form is similar to the modern tufas described by Jones (1925). The second type is porous and laminated, develops regular, dome-shaped structures. (See pl. 22, fig. 2, and pl. 24, fig. 7.) This type of tufa resembles the algal structures of Andros Island and the Belt series. (See fig. 52; pl. 20, fig. 3, and pl. 22, fig. 3.) I believe that these structures might have developed above the lake level and that the lamination is due to alteration of the paperlike mat, described by Jones, and very fine detrital sediment.
The occurrences described in the foregoing discussions are outlined below according to the environments and the types of spongiostromes found in the environments. Paleoclimatic data from Lake Lahonton was furnished by Roger Morrison (written communication, 1954).
Environments in which spongiostromes form
| I. Marine environments. | ||||
| A. Shallow marine environments, depths of 1-6 feet. | ||||
| 1. Oncolites (unattached spheroidal bodies). | ||||
| B. Environments that are near sea level but above low watermark. | ||||
| 1. Sediment-binding algae that may produce stromatolites similar to some of those of the Belt series. (See p. 141.) | ||||
| C. Deeper marine environments. | ||||
| 1. Calcareous algae (red or green algae) exist at greater depths but these are not considered to be spongiostromes. | ||||
| II. Continental environments. | ||||
| A. Fresh-water lakes, such as Green Lake, N. Y., in which the algal deposits develop below the surface of the water. (See p. 144.) | ||||
| 1. Structures that are predominantly arborescent rather than stromatolitic. | ||||
| 2. Concretionary structures (water biscuits) that should be called oncolites rather than stromatolites. | ||||
| B. Fresh-water streams, such as Furnace Creek, W. Va. (See p. 144.) | ||||
| 1. Predominantly concretionary structures (oncolites). | ||||
| C. Lakes of the Great Basin section, such as Lakes Lahonton and Bonneville and their remnants, in which salinity varies appreciably with time. (See p. 145.) | ||||
| 1. Shallow lakes, saline and warm, with fluctuating lake level and generally slight wave action. | ||||
| a. Beach zones (areas kept wet by wave action and extending several feet above lake level at a given time). | ||||
| (1) Tufa, porous, laminated, lithoid; closely resembles the stromatolites of the Belt series. | ||||
| b. Lake bottoms, shallow water. | ||||
| (1) Tufa deposits, compact, laminated, irregularly shaped, lithoid. These forms only crudely resemble the stromatolites of the Belt series. | ||||
| 2. Deep lakes, relatively fresh cold water; fluctuating lake level; strong to weak wave action at times of tufa deposition. | ||||
| a. Beach zones. | ||||
| (1) Forms similar to IIC1a(1), above. | ||||
| b. Lake bottoms, shallow to deep water. | ||||
| (1) Arborescent forms similar to deposits of modern fresh-water lakes. (See IIA1, above.) | ||||
The similarities between some of the lithoid tufas of Lake Lahonton and the algal deposits of Andros Island is not a surprising coincidence. Both environments are close to the level of a standing body of water and in both places the algae develop in a zone of extreme variability in salinity. The hurricane winds that occasionally drive waves across the interior of Andros Island have their counterpart in the Great Basin section. Morrison (oral communication, 1954,) cites occasions when strong winds blowing across the Carson Desert (a huge playa about 20 miles in diameter) drive water from the Carson Sink into areas that normally would be considerable distances from the shore of the lake.
ORIGIN OF STROMATOLITES
Previous workers have regarded the Cyanophyta and possibly the unicellular Chlorophyta as the organisms responsible for stromatolites. The foregoing discussions seem to substantiate this view. However, several important environmental limiting conditions have not been exhaustively explored in considering fossil forms.
The first limiting factor is the depth of water in which the stromatolites developed. Most authors state simply that they developed in shallow water and refer to the maximum penetration of light as the most important factor in controlling the distribution of the Cyanophyta. Cloud (1942, p. 371) reports that the transparency of fresh water is much less than that of the sea water and that stromatolite-building blue-green algae probably do not form large structures below about 10 meters in fresh water and 30 meters in the sea. I am convinced, from my own observations and those of others, that the Cyanophyta are dominant only above low watermark in the marine environment. Modern stromatolites are found only above low watermark. Black's type A grows in the intertidal zone near high watermark. As the tides on Andros Island range from 2.3 feet to 3.0 feet (U. S. Navy, 1951, p. 67), the depth of water over the algal deposits when submerged must be very slight indeed. Here we encounter one of the difficulties in comparing living and fossil stromatolites. Recent stromatolites are quite small, being generally less than a foot in diameter. Many fossil forms are of comparable size or somewhat larger, but a few range up to several feet in diameter and up to 10 feet in height.
Ginsburg (written communication, 1954) seems to think that the difference in scale is not important. His arguments are based primarily upon structural similarity. Cloud, however, (written communication, 1954) feels that during Belt time sea tides of improbably great amplitude (about 10 feet) would have been required to account for such large structures. He prefers the theory that these structures grew below low watermark—on the basis that the blue-green algae have had a long time for habitat variations since the Paleozoic and that perhaps they were forced out of the ocean by competition from better adapted forms. This statement is very possibly true. However, in the light of my observation of algal mats growing on the bottoms of fresh-water lakes, I cannot conceive of a process that would form headlike structures similar to stromatolites other than periodic exposure to subaerial conditions. In all the fresh-water lakes that I have found algal mats, the mats have smooth surfaces that conform to the lake bottoms. Admittedly, the mats will cover irregularities on the bottom, but there is a tendency for the mats to decrease the irregularities rather than intensify them.
A second possible limiting condition involves the salinity of the water in which the stromatolites develop. There has been some disagreement over the question of a marine origin vs. a fresh-water origin of these structures. According to Black, the stromatolites of the Bahama Islands range from fresh lake water to water of ocean salinity. The range of salinity is even greater in the continental environments. Here, the environments range from lakes of fresh water, as in central New York, to lakes such as Lake Lahonton, whose salinity has varied greatly with climatic changes and the resultant fluctuations in lake volume during the late Quaternary. It seems safe to state at this time that fossil stromatolites can give us little aid in determining the salinity of ancient lakes.
The third condition is of extreme importance. It concerns the manner in which stromatolites grow. Do they grow by the precipitation of lime from water as a result of photosynthesis, or do the algae develop on the surfaces of unconsolidated, stratified sediments and merely bind the sediments together. Some algae cause the precipitation of lime—either as a loose precipitate or as a hard, stony incrustation around the algal colonies. Several occurrences of this type have been reported in the published record. Probably the best known is the one at Green Lake, N. Y. Here, the form of the deposit is controlled by the growth habit of the algae, and it can be readily seen that a fine precipitate of lime is deposited about the algal filaments. This deposit of finely divided lime adheres to the mucilaginous sheaths of the algae and is gradually built up into a spongy porous mass. Roger Morrison states (oral communication, 1954) that nowhere in the Lahonton region are arborescent algal deposits growing today. Presumably, the highly alkaline composition of the water does not permit the growth of species that develop this form.
The algal mats of Andros Island and the Carson Sink, on the other hand, do not add lime to the sediments. The sediments on Andros Island are fine sands and carbonate muds. The algae grow on the surfaces of these sediments; the filaments permeate the sediment as it is deposited and play an important part in binding it. This mode of stromatolite origin has been suggested by Black (1933, p. 167, 186; 1938, p. 166), Young (1935, p. 158), and Cloud and Barnes (1948, p. 98, 100). Some of the stromatolites of the Belt series are believed to have developed in this manner. Structures such as Collenia frequens Walcott probably did not grow above the surface of deposition as unsupported columns, but developed gradually as the surface was raised due to sedimentation. The columns developed gradually by the periodic increment of algal layers similar to the ones that make up Black's type B.
PALEOECOLOGY
COLLENIA FREQUENS ZONE
It is possible that this zone was formed under conditions similar to those on Andros Island, where Black's type B occurs. The sediment is a fine-grained dolomitic limestone that is interbedded with edgewise breccia and lenses of dolomitic sandstone or conglomerate. These structures are indicative of frequent disturbance by agitated waters, such as may occur on tidal flats.
Thin sections show an alternation of a fine carbonate mosaic with layers of coarse calcite and dolomite (similar to specimens shown in pl. 24, figs. 5 and 6) The grains in the mosaic average about 4 microns in diameter. Conceivably, this fine-grained carbonate was a chemical precipitate mechanically entrapped about the filaments of the algae. The coarsely crystalline calcite and dolomite represent the detrital material that was deposited over the surface of the algal mat during times of high water and considerable agitation.
NEWLANDIA LAMELLOSA WALCOTT
This form suggests development on mud flats that were alternately wet and dry. Dessication would have caused the edges of the colonies to curl up so as to form bowl-shaped structures. (See pl. 23, fig. 8; and fig. 55.) After dessication of the original crusts, new algal mats formed within the bowl-shaped structures, and these were less concave than the original.
COLLENIA UNDOSA ZONES
Collenia undosa Walcott does not conform to any of the living types described by Black. Wherever this species has been observed, its mineralogical composition differs greatly from the surrounding sediments. The stromatolite invariably occurs as alternating layers of argillite and pure calcite. The enclosing sediment is either argillite or coarse calcareous sandstone. The form of the calcite laminae crudely resembles that of some of the modern encrusting coralline algae.
This species is ordinarily associated with Cryptozoon occidentale Dawson and Collenia symmetrica Fenton and Fenton and is found only in the Collenia undosa zones. The colonies are generally noted to have grown on mud-cracked surfaces. No evidence for desiccation is found after the inception of the colonies, and so it is assumed that they developed upon subsiding surfaces. The presence of coarse limesand and conglomerate, consisting of stromatolite fragments, surrounding the colonies suggests to me that they were subjected to wave action throughout their growth.
COLLENIA SYMMETRICA ZONES
A comparison of figure 54 with figure 5 of plate 22 shows the resemblance between Collenia symmetrica Fenton and Fenton and Black's type C. The one difference between the two forms is that most colonies of Collenia symmetrica do not indicate a period of desiccation after their inception. This species grew upon mud-cracked surfaces which influenced the shape of the laminae in the younger portion of the colonies. Later they formed dome-shaped cross sections. In some of the outcrops, edgewise breccia of stromatolite fragments overlies the biostromes of Collenia symmetrica. (See pl. 22, fig. 5.) The lack of evidence for desiccation after the inception of the colonies and the evidence for strong agitation suggests that this species may have formed upon subsiding mud fiats.
CONOPHYTON ZONES
Conophyton inclinatum n. sp. is the only species that appears to have formed below the zone of agitation. The species is restricted to the two Conophyton zones and seems to have grown under a special set of conditions. No other species of stromatolite is known to occur in the same beds with Conophyton. There is no evidence for desiccation or fragmentation of the colonies by wave action. The surrounding sediments consist of finely laminated limestone that presumably was deposited in a quiet environment. The direction of growth of the colonies is inferred to have been controlled by current action inasmuch as the long axes of the cones lie at low angles to the bedding surfaces.
The colonies themselves are different from other species in the area. They appear to have been attached to the substratum by their conical basal termini. Growth was by the addition of cone-shaped laminae, one upon the other. No forms similar to Conophyton are known to be growing at the present time.
The succession of biostromes in the Conophyton zone 1 indicates a gradual change from a mud-flat environment, on which Collenia frequens Walcott developed, to an offshore or possibly lagoonal environment, in which Conophyton grew, to finally a mud-flat environment, on which Cryptozoon occidentale Dawson and Collenia multiflabella n. sp. were the dominant forms.
In the Conophyton zone 2 the succession is interpreted to represent an alternation of offshore and mud-flat environments. Two cycles are indicated by the alternation of biostromes of Conophyton inclinatum n. sp. and Collenia frequens Walcott. (See p. 140.) Subzone A presumably formed as the result of the recession of waters in which the underlying limestones were deposited. On the new mud flats Collenia frequens Walcott grew to a thickness of 23 feet. A renewed advance of the shoreline, which caused the mud flats to be permanently submerged, may have fostered growth of colonies of Conophyton inclinatum n. sp. Conditions on the sea bottom then presumably remained the same until a thickness of 35 feet of Conophyton had accumulated.
Some event, perhaps fouling of the water, created conditions that were unfavorable for the growth of Conophyton, and 11 feet of black shaly limestone was deposited. Conophyton may well have been growing in an embayment that received a plentiful supply of well-oxygenated water from the Belt sea. If the embayment were isolated from the sea, the dissolved oxygen near the bottom would have become exhausted and the oxidizing environment would then have given way to a reducing environment in which only anaerobic bacteria could survive. The deposits of such an environment are ordinarily black muds (Hatch, Rastall, and Black, 1938, p. 275). It is likely that subzone C was deposited in such an environment. Filling of the embayment with sediment produced a mud flat and the renewed growth of Collenia frequens Walcott. This condition did not last long, for only about 6 feet of Collenia frequens Walcott (subzone D) was formed. Another advance of the shoreline brought back the conditions under which subzone B was deposited, and 24 feet of Conophyton beds was deposited to form subzone E. The final event was possibly further deepening, which caused the cessation of the growth of Conophyton and the deposition of barren limestone.
COLLENIA MULTIFLABELLA ZONE
This zone represents the continuation of the trend that was begun during the latter part of the Conophyton zone 1. This trend consisted of the beginning of the deposition of argillaceous sediments and probably shallowing of the sea. The amount of argillaceous material increases upward. The boundary between the Siyeh limestone and the Missoula group is placed at the top of this zone. Above lie the argillites of the lower part of the Missoula group.
The dominant species is Collenia multiflabella n. sp. Subordinate amounts of Cryptozoon occidentale Dawson also occur. The basal portions of Collenia multiflabella closely resemble Black's type B and may have formed under somewhat similar conditions. A slight change in the environment, perhaps an influx of finely divided sediment, caused the filling in of the low spots between the algal heads. The new algal mats that grew upon the new surface were large enough to cover several of the older heads.
| <<< Previous | <<< Contents >>> | Next >>> |
pp/294-D/sec4.htm
Last Updated: 18-Jul-2008